Гіпоталамус Нейроендокринологія секреції ГнРГ
Гіпоталамічний контроль секреції гонадотропінів гіпофізом здійснюється за допомогою гонадотропін-рилізинг гормону (ГнРГ).
Утворення ГнРГ-декапептиду кодується геном, локалізованим на короткому плечі хромосоми 8.
ГнРГ-рецептори локалізуються в гіпофізі та багатьох органах репродуктивної системи. Нейрони, які секретують ГнРГ, знаходяться переважно у передньому, медіобазальному (горбовому) гіпоталамусі та аркуатних ядрах. По аксонах цих нейронів ГнРГ транспортується до медіального підвищення гіпофіза, де досягає капілярів первинного портального сплетення. Нервові клітини, які транспортують ГнРГ з аркуатних ядер гіпоталамуса до медіального підвищення гіпофіза, одержали назву горбо-лійкового, або горбо-гіпофізарного шляху.Медіальне підвищення, або лійка, разом з лійковим стеблом і задньою часткою гіпофіза утворюють нейрогіпофіз. Ці три компоненти нейрогіпофіза утворюють загальну капілярну сітку і мають пряме артеріальне кровопостачання з гіпофізарних артерій. Капіляри медіального підвищення мають фенестрований епітелій, як і периферичні тканини, що дозволяє пропускати великі молекули. Ці капіляри відрізняються від капілярів головного мозку, отже медіальне підвищення гіпофіза знаходиться поза гематоенцефалічним бар’єром.
Нервові клітини з горбо-гіпофізарного тракту секретують ГнРГ безпосередньо у портальну циркуляцію, звідки він переноситься в клітини передньої частки гіпофіза. Медіобазальна частина передньої частки гіпофіза (аденогіпофіз) одержує кров з портальних судин гіпофіза і локалізується близько до основи гіпоталамуса і гіпофі- зарного стебла (рис. 4.1). На відміну від нейрогіпофіза, аденогіпофіз не має прямого
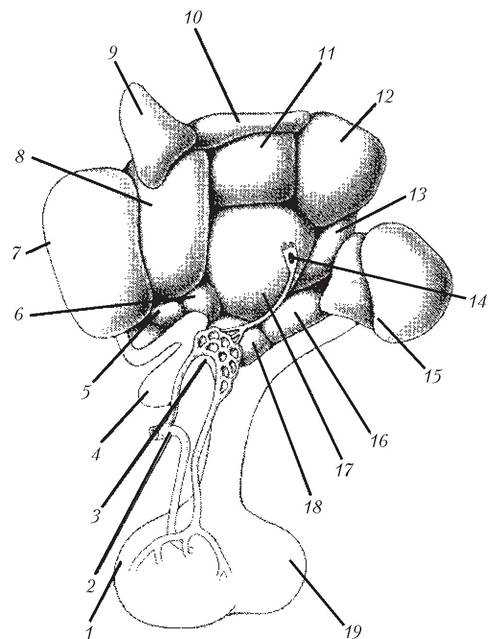
Рис. 4.1. Ядерна структура гіпоталамуса (сагітальний розріз у площині III шлуночка мозку; каудальна частина праворуч):
1 — передня частка гіпофіза; 2 — верхня гіпофізарна артерія; 3 — медіальне підвищення; 4 —перехрестя зорових нервів (хіазма); 5 — супрахіаз- матизні ядра; 6 — супраоп- тичні ядра ; 7 — преоптичні ядра; 8 — передня зона гіпоталамуса; 9 — паравентрикуляр- не ядро; 10 — дорсальна зона гіпоталамуса; 11 — дорсомеді- альне ядро; 12 — заднє гіпота- ламічне ядро; 13 — премамі- лярне ядро; 14 — таніцит; 15 — мамілярне тіло; 16 — горбова зона; 17 — вентромедіальне ядро; 18 — аркуатне ядро; 19 — задня частка гіпофіза
артеріального кровопостачання і одержує кров з портальних судин.
Залишаючи аде- ногіпофіз, судини повертаються до нейрогіпофізарного капілярного сплетення, дозволяючи гіпофізарним гормонам брати участь у регуляції секреції ГнРГ медіальним підвищенням.Додатково до цього основного шляху існують альтернативні шляхи транспорту ГнРГ. Аксони горбо-лійкового тракту можуть переносити ГнРГ прямо до третього шлуночка мозку. Спеціалізовані епендимальні клітини — таніцити — поширюються з порожнини третього шлуночка в найдальшу зону медіального підвищення (рис. 4.2). Якщо ГнРГ потрапляє у третій шлуночок, він транспортується у портальну систему через мікроворсинки таніцитів. Отже, ГнРГ виділяється двома шляхами: у великій кількості періодично через горбо-лійковий тракт (циклічне виділення) і у незначній кількості постійно через трансепендимальну систему (тонічне виділення).
У жінки ГнРГ секретується в пульсаційному режимі і має період напівжиття 24 хв. Амплітуда і частота імпульсів варіює протягом менструального циклу, частота є більшою у фолікулярну фазу (1 імпульс за 1 год) і меншою у лютеїнову фазу (1 імпульс за 2-3 год). Пульсація ГнРГ взаємодіє з секрецією гонадотропінів. Так, при щогодинному екзогенному введенні ГнРГ виникає пік секреції гонадотропінів, характерний для овуляторної фази менструального циклу, а при введенні ГнРГ з частотою 5 імпульсів за годину секреція гонадотропінів пригнічується. При зменшенні частоти введення ГнРГ до 3 імпульсів за годину рівень лютеїнізуючого гормо-
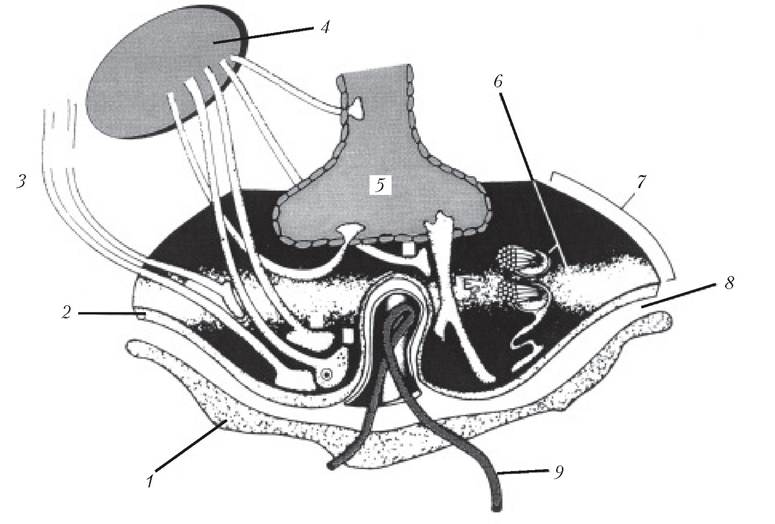
Рис. 4.2. Схематична діаграма таніцитів (8), які проходять між третім шлуночком і зовнішньою частиною медіального підвищення гіпоталамуса:
1 — горбова частина; 2 — базальна мембрана; 3 — нейро-ендокринні клітини (шляхи виділення серотоніну); 4 — горбо-лійковий (тубероінфундибулярний) тракт; 5 — третій шлуночок; 6 — аксони супраоптичних і паравентрикулярних нейронів; 7 — медіальне підвищення; 8 — периваскулярний простір; 9 — портальні судини
ну (ЛГ), або лютропіну, зменшується, а рівень фолікулостимулювального гормону (ФСГ), або фолітропіну, зростає; не виникає овуляторного піка секреції гонадотро- пінів. Зменшення кількості екзогенного ГнРГ також пригнічує виділення гонадотро- пінів гіпофізом.
Жінки з ановуляцією та гіпоталамічною аменореєю, а також з аменореєю нез’ясо- ваного походження мають порушення частоти та (або) амплітуди виділення ГнРГ. Отже, контроль епізодичної секреції ГнРГ є дуже важливим для підтримання нормальної овуляторної циклічності. Амплітуда і частота ГнРГ-секреції гіпоталамусом регулюється зворотним зв’язком як з яєчниковими стероїдами — естрадіолом і прогестероном, так і з гіпофізарними гонадотропінами гуморальним шляхом. Амплітуда і частота імпульсів ГнРГ регулюється також неврогенним шляхом за допомогою ней- ротрансмітерів і нейромодуляторів головного мозку.