Тришаровий зародковий диск. Гаструляція
Найхарактернішою подією третього тижня розвитку є гаструляція — утворення трьох зародкових листків — ектодерми, мезодерми і ендодерми ембріона (рис. 5.9). Гаструляція починається з утворення первинної смужки на поверхні епібласта.
Головний кінець смужки — первинний вузлик — локалізується навколо первинної ямки. У ділянці первинного вузлика та первинної смужки клітини епібласта зміщуються досередини (інвагінують), утворюючи два нових клітинних шари — ендодерму і мезодерму. Клітини, які залишаються у складі епібласта, утворюють ектодерму. Отже, епібласт дає початок усім трьом зародковим листкам ембріона: ендодермі, мезодермі і ектодермі. Клітини внутрішньозародкової мезодерми мігрують доти, доки не зустрінуться з позазародковою мезодермою, яка вкриває жовтковий мішок і амніон. У головному напрямку вони розходяться по обидва боки від прехордіальної пластинки. Остання формується між верхівкою нотохондри та щічно-глотковою мембраною і є важливим фактором індукції розвитку переднього мозку.Пренотохондріальні клітини інвагінують у первинну ямку, рухаючись у краніальному напрямку і досягають прехордіальної пластинки. Ці пренотохондріальні клітини вселяються в гіпобласт таким чином, що згодом серединна лінія ембріона складається з двох шарів клітин, які формують нотохордіальну пластинку. У міру заміщення гіпобласта клітинами ентодерми, які вростають усередину ембріона в ділянці первинної смужки, клітини нотохордіальної пластинки проліферують та відділяються від ентодерми і згодом утворюють суцільний тяж клітин — дефінітивну ното- хорду, що лежить під нервовою трубкою і є зачатком осьового скелета. Спочатку формується краніальний кінець, потім первинна смужка поширюється у каудальному напрямку і утворює каудальний кінець ембріона. У місці, де ямка утворює заглибину в епібласті, утворюється нейрокишковий канал, що тимчасово сполучає порожнини амніона і жовткового мішка.
На каудальному кінці ембріона з щільно зрощених клітин ектодерми і ентодерми, без проміжного шару мезодерми, утворюється клоакальна мембрана. Після появи клоакальної мембрани, близько 16-го дня розвитку, задня ділянка стінки жовткового мішка формує невеликий дивертикул, який вростає у сполучну ніжку — алантокишковий дивертикул, або алантоїс. У людини алантоїс залишається рудиментарним і може спричинювати вади розвитку сечового міхура.Перед початком і в процесі гаструляції відбувається становлення осей тіла: пере- дньозадньої, дорзовентральної і лівоправосторонньої. Клітини задньої крайової зони (PMZ) на каудальному кінці ембріона виділяють активінподібну речовину сімейства генів, трансформуючих фактори росту β (TGF-β), яка індукує утворення первинної смужки. Цим визначається краніально-каудальна вісь зародка. Після утворення первинної смужки низка генів регулює формування дорзальної та вентраль-
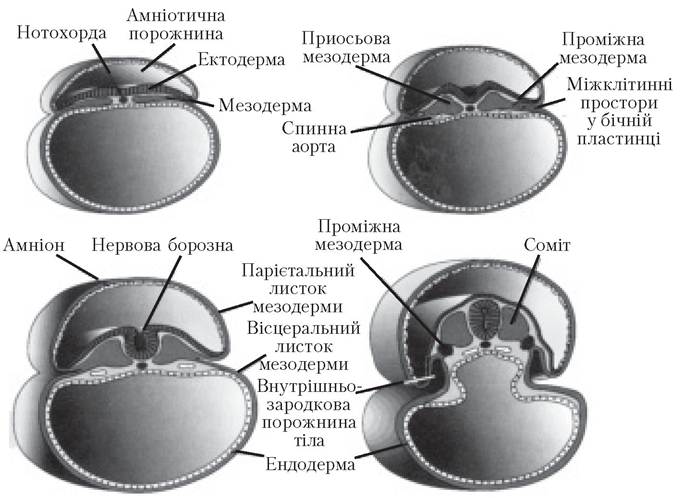
Рис. 5.9. Розвиток мезодермального зародкового листка (17-та-21-ша доба)
ної мезодерми, а також головних і хвостових структур. Інший ген сімейства TGF-β, кістковий морфогенетичний протеїн-4 (ВМР-4), продукується на всій площі ембріонального диска. У присутності цього протеїну та фактора росту фібробластів (FGF) мезодерма вентралізується, забезпечуючи утворення нирок (проміжна мезодерма), крові, а також мезодерми стінки тіла (мезодерма бічної пластинки). Хордин, ногін і фолістатин пригнічують активність ВМР-4 і забезпечують дорзалізацію мезодерми, внаслідок чого утворюються нотохорда і сомітомери ділянки голови. За відсутності фактора HNF-3β ембріони не здатні до нормальної гаструляції та не мають переднього та середнього мозку. Ген Goosecoid активує інгібітори ВМР-4 і регулює розвиток голови. Надлишкова або недостатня експресія цього гена призводить до тяжких вад розвитку голови, включаючи її подвоєння.
Утворення дорзальної мезодерми в серединній і каудальній ділянках-регіонах регулює ген Brachyury (Т).
Відсутність цього гена спричинює вкорочення ембріональної осі — каудальний дисгенез. Ступінь вкорочення залежить від терміну виникнення дефіциту цього протеїну.Встановлення лівоправосторонньої асиметрії регулюється каскадом генів, у тому числі sonic hedgehog (Shh), продукт активності якого надходить до лівої частини первинного вузлика за допомогою рецептора активіну-ІІа, експресію якого здійснює права частина первинного вузлика. Протеїн Shh активує гени nodal і lefty, які проявляються у мезодермі бічної пластинки лівої сторони зародка і регулюють активність інших генів для забезпечення асиметрії так, що серце і селезінка опиняються у лівій половині тіла, а більша частина печінки — у правій.
Клітини епібласта, які мігрують через первинний вузлик і смужку, предетерміно- вані своїм місцем знаходження до утворення певних типів мезодерми і ендодерми. Так, клітини, що вростають через краніальну ділянку первинного вузлика, стають нотохордою, ті, що мігрують через латеральні краї первинного вузлика і краніальний кінець первинної смужки, стають приосьовою мезодермою. Клітини, які мігрують через серединну ділянку первинної смужки, стають проміжною мезодермою, а ті, що мігрують через каудальну частину первинної смужки, формують мезодерму бічної пластинки. Отже, первинна смужка забезпечує краніокаудальну організацію зародкової мезодерми.
Ембріональний диск, на початку плоский і майже круглий, поступово стає продовгуватим з широким головним (цефальним) і вузьким хвостовим (каудальним) кінцем. Розширення ембріонального диска відбувається переважно у головному кінці, що спричинено постійною міграцією клітин з первинної смужки у головному напрямку. Інвагінація поверхневих клітин первинної смужки та їх подальша міграція вперед і латерально спостерігаються до кінця четвертого тижня. Далі первинна смужка регресує. Отже, у головному кінці зародкові листки починають специфічну диференціацію у середині третього тижня, тимчасом як у хвостовому кінці така диференціація починається наприкінці четвертого тижня.
Отже, гаструляція, або формування зародкових листків, продовжується у каудальних сегментах, тимчасом як краніальні структури диференціюються і обумовлюють цефалокаудальний розвиток ембріона.Клінічні кореляції. Гаструляція — критичний період розвитку ембріона. Початок третього тижня розвитку (через чотири тижні від останньої менструації, або через два тижні після запліднення), коли відбувається гаструляція, є надзвичайно чутливим періодом щодо тератогенних впливів. У цей час відбувається закладення мозку, очей та інших тканин, які можуть бути ушкоджені дією агресивних факторів-тера- тогенів. Так, наприклад, дія великих доз алкоголю в цей період призводить до деградації клітин переднього кінця серединної лінії диска, внаслідок чого розвивається недостатність краніофасціальних структур і голопрозенцефалія. Голопрозенцефалія характеризується зменшенням переднього мозку, має місце злиття латеральних шлуночків мозку, а також гіпотелоризм (очі розміщені близько одне до одного).
Каудальний дисгенез (сиреномелія) — синдром, який полягає у недостатності утворення мезодерми у крайньому каудальному відділі ембріона. З цієї мезодерми формуються нижні кінцівки, сечостатева система (проміжна мезодерма) та попереково- крижові хребці, мають місце аномалії цих структур: гіпоплазія або злиття нижніх кінцівок, аномалії хребців, агенезія нирок, атрезія ануса, аномалії статевих органів. Такі аномалії розвитку асоціюються з декомпенсованим цукровим діабетом у матері.
Situs inversus (обернене положення) — стан, при якому має місце транспозиція органів грудної клітки і черевної порожнини. У 20 % таких хворих наявні бронхоектазії і хронічні синусити, спричинені аномаліями війок (синдром Картагенера). Війки у нормі присутні на вентральній поверхні первинного вузлика і можуть розподілятися по різних структурах організму під час гаструляції. Інший приклад аномалій позиції відомий під назвою латеральних послідовностей. Хворі з такими вадами не мають повного situs invertus, але виявляються переважно білатерально ліво- або пра- восторонніми. Так, для лівосторонньої білатеральності притаманна поліспленія, для правосторонньої — аспленія або гіпоплазія селезінки. У хворих із латеральними послідовностями виявляються й інші аномалії, особливо серця.
Пухлини, асоційовані з гаструляцією. Якщо залишки первинної смужки персисту- ють у крижово-куприковій ділянці, вони можуть проліферувати і утворювати пухлини, відомі як крижово-куприкові тератоми. Ці пухлини нерідко походять з усіх трьох зародкових листків і є найбільш частими пухлинами у новонароджених (1:37 000).