Транспорт холестерина в субэндотелий
Основным переносчиком холестерина (его в мицелле этого вида липопротеинов содержится более 50%) являются ЛИНИ В ЛПОНП холестерина содержится несколько меньше. Этот вид липопротеинов в основном является переносчиком триглицеридов.
ЛПВП содержат очень мало холестерина и много белка и фосфолипидов.JШНП доставляют холестерин к клеткам организма, в том числе - и к клеткам эндотелия сосудов. ЛПВП, наоборот, переносят холестерин от клеток различных органов и тканей к клеткам печени, где он и метаболизируется. Следует иметь в виду, что холестерин может проникнуть в клетку только благодаря рецепторам JІНШІ. При соединении ЛПНП с соответствующими рецепторами (рецепторы ЛПНП), холестерин выделяется из мицеллы, переносится в протоплазму клетки и там используется для ее нужд. С другой стороны, при переносе холестерина из клеток организма (в том числе - и из клеток сосудистого эндотелия) к клеткам печени с помощью ЛПВП он может проникнуть в гепатоциты только благодаря рецепторам ЛПВП, находящимся на мембране гепатоцитов. Таким образом, как усвоение холестерина клетками эндотелия сосудов, так и перенос излишков холестерина от клеток эндотелия сосудов к гепатоцитам и усвоение ими холестерина зависит от состояния и плотности соответствующих рецепторов на мембране клеток. В норме клетки способны увеличивать или уменьшать количество рецепторов как ЛПНП, так и ЛПВП. Однако, в значительной степени эта их особенность генетически детерминирована.
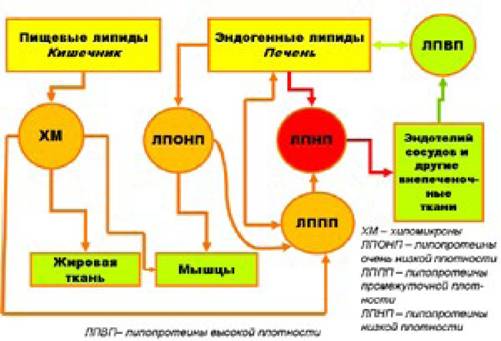
Схема осуществления транспортной функции липопротеинов
Весьма серьезную роль в контактах липопротеинов с рецепторным аппаратом клеток играют апопротеины, которые, как мы помним, встроены в липидный монослой мембраны липопротеиновой мицеллы. Именно молекулы апопротеинов мицеллы липопротеинов являются лигандами, связывающими соответствующий липопротеин с его рецептором.
Они же в ряде случаев играют и роль кофакторов для соответствующих ферментов. В настоящее время выделено четыре вида апопротеинов.Апо-А - обеспечивает связь ЛПВП с соответствующим рецепторным аппаратом.
Апо-В - обеспечивает связь ЛПНП с рецепторным аппаратом клеток печени и периферических тканей;
Апо-С-II - кофактор для липопротеиновой липазы, благодаря которой триглицериды удаляются из хиломикронов и ЛПОНП;
Апо-Е - обеспечивает связь липротеинов с рецепторным аппаратом гепатоцитов.
Описан и еще один вид апопротеинов - так называемый апопротеин (а), которому приписывают наибольшую атерогенность и входящий в состав ЛИНИ. Атерогенность этого апопротеина объясняют несколькими его свойствами. Во-первых, он легко окисляется и поглощается макрофагами. Во-вторых, печеночные клетки имеют наименьшее количество рецепторов к ЛИНИ, содержащим липопротеин (а). И, наконец, в-третьих, имеются данные о том, что ЛИНИ, содержащие липопротеин (а), обладают повышенными антитромболитическими свойствами.
Учитывая особенности транспорта холестерина липопротеинами (от печени к клеткам других органов и тканей и от клеток органов и тканей к клеткам печени) одни из них (ЛИОНИ и, особенно, ЛИНИ) получили название «атерогенных липопротеинов», а другие (ЛПВП) - «антиатерогенных липопротеинов». Однако, говорить о том, что патогенез атеросклероза определяется только уровнем концентраций в плазме крови атерогенных и антиатерогенных липопротеинов, конечно, нельзя. Нужно не забывать о достаточности (недостаточности) соответствующего рецепторного аппарата, а также о полноценной (или неполноценной) функции апопротеинов, входящих в состав мицеллы липротеинов. Кроме того, значительную роль в провокации атеросклероза играют и, так называемые, «модифицированные» липопротеины. Иричин модификации (изменения) липопротеинов достаточно много[2], однако основной из них является их перекисное окисление. В результате перекисного окисления «нормальные» липопротеины приобретают атерогенные свойства, которые объясняются двумя
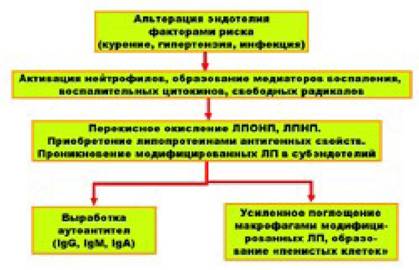
Перекисное окисление ЛПНП и образование «пенистых» клеток
причинами.
Во-первых, модифицированные липопротеины могут без каких-либо ограничений накапливаться в макрофагах, что заставляет их превращаться в «пенистые клетки» - один из основных факторов морфогенеза атеросклероза. Известно, что макрофаги в норме поглощают избыток ЛПНП и, метаболизируя их, предотвращают перегрузку клеток эндотелия сосудов холестерином. Этот процесс регулируется по принципу обратной связи: при заполнении макрофага достаточным количеством ЛПНП чувствительность рецепторного аппарата макрофага к ЛПНП резко падает и он перестает их поглощать. Модифицированные ЛПНП нарушают эту регуляционную связь, и макрофаги поглощают эти липотротеины без каких-либо ограничений, вплоть до их полной перегрузки и превращения в пенистые клетки.
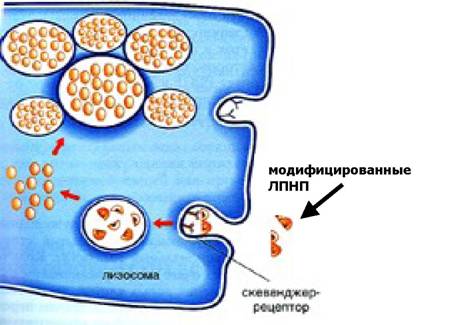
Захват модифицированных ЛПНП скевенджер-рецепторами макрофагов (по О.П.Шевченко, О.Д.Мишнёв)
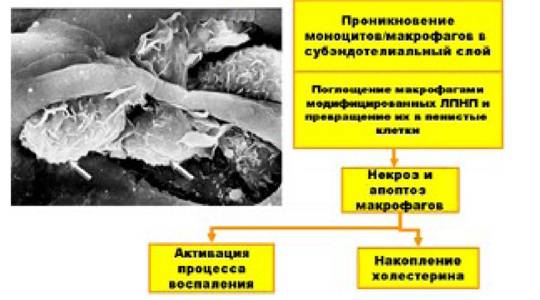
Роль макрофагов, трансформирующихся в пенистые клетки, в развитии атеросклероза
Важную роль в захвате модифицированных ЛПНП макрофагами играют, так называемые, рецепторы-мусорщики (скевенджер-рецепторы), которые синтезируются в макрофагах, перегруженных модифицированными ЛПНП. Именно образование этих рецепторов и нарушает деятельность обратной регулирующей связи, которая была уже упомянута выше.
Во-вторых, белки, входящие в состав модифицированных липопротеинов, приобретают антигенные свойства, благодаря чему к ним начинается выработка аутоантител, повреждающих, в том числе, и клетки эндотелия сосудов и тромбоциты. Так запускается еще один патологический цикл, лежащий в основе развития атеросклероза.
Таким образом, об «атерогенности» и «антиатерогенности» тех или иных липопротеинов, если мы имеем в виду нормальный организм, говорить не приходится. «Атерогенность», например, ЛПНП проявляется только тогда, когда в действие вступают другие факторы патогенеза атеросклероза.
7.