Дія гормонів на специфічні репродуктивні фактори Яєчниковий цикл (яєчниковий гаметогенез, оогенез)
Навіть без стимуляції гонадотропінами примордіальні фолікули можуть розвиватись у первинні (преантральні) фолікули. Наявність ФСГ в овуляторному циклі призводить до зростання кількості гранульозних клітин навколо ооцита, дозрівання фолікула до антральної стадії й утворення центральної порожнини — фолікулярної печери (antrum).
Без ФСГ-стимуляції всі фолікули піддаються атрезії. Циклічне дозрівання первинних фолікулів звичайно починається у періоді статевого дозрівання. Сполучна дія ФСГ і ЛГ сприяє зростанню виробки естрадіолу гранульозними клітинами. Естрадіол стимулює ріст преантральних фолікулів, запобігає їх атрезії і посилює дію ФСГ на гранульозні клітини. Тестостерон сприяє атрезії фолікулів і запобігає росту преантральних фолікулів. Місцевий вміст естрогенів та андрогенів у фолікулі впливає на шлях його розвитку: дозрівання або атрезія. Домінантний фолікул секретує велику кількість естрадіолу, що сприяє збільшенню кількості рецепторів ФСГ, зростанню кількості гранульозних клітин та їх мітотичної активності.Підвищення рівня естрадіолу викликає ефект негативного зворотного зв’язку щодо виділення ФСГ гіпофізом і розвиток інших фолікулів припиняється (атрезія). Гранульозні клітини домінантного фолікула секретують інгібін, який також пригнічує виділення ФСГ. Ооцит оточується прозорою оболонкою (zona pellucida) та променистим вінцем (corona radiata), у фолікулі накопичується рідина, яка містить естрогени, андрогени та протеїни. Після досягнення критичної маси гранульозних клітин у фолікулі виникає печера (antrum) і ооцит переміщується в бік фолікула (рис.4.7). За даними ультразвукового дослідження, домінантний фолікул досягає діаметра в середньому 19 мм (18-25 мм), а його об’єм зростає до 3,8 мл (3,1-8,2 мл). Близько ооцита гранульозні клітини нагромаджуються і утворюють яйценосний горбок (cumulus oophorus). Під час проліферації гранульозних клітин на їх поверхні утворюються рецептори до ЛГ.
При зв’язуванні молекул ЛГ зі специфічними рецепторами проліферація гранульозних клітин припиняється і вони починають секретува- ти прогестерон.Близько 80 % усієї кількості естрадіолу (500 мкг) продукується домінантним фолікулом напередодні овуляції ( за 12-24 год). Зростання рівня естрадіолу і, в меншій мірі, прогестерону стимулює секрецію ЛГ гіпофізом (позитивний зворотний зв’язок). Пік ЛГ у середині циклу ініціює процес овуляції — розриву стінки зрілого фолікула і виходу ооцита в черевну порожнину і в подальшому в просвіт маткової труби. Овуляція виникає приблизно через 24 год після досягнення піка секреції
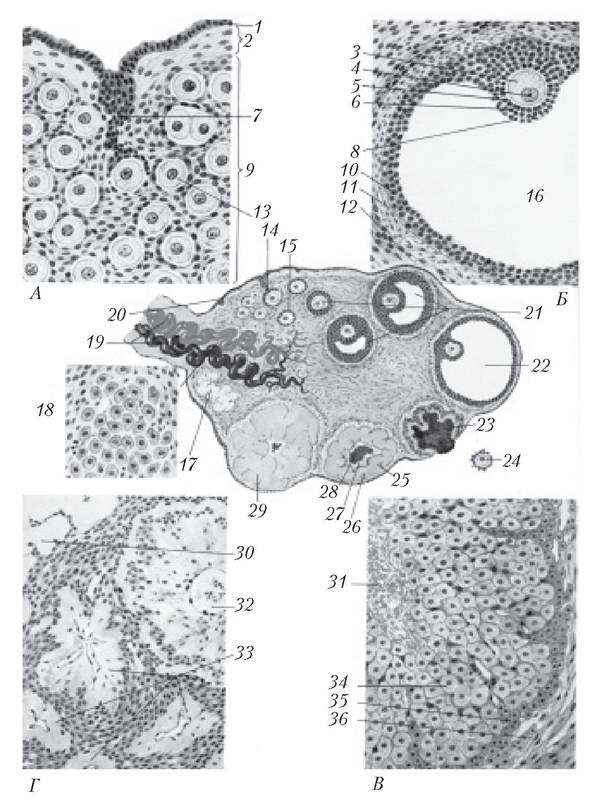
Рис. 4.7. Анатомія і фізіологія яєчника (А — яєчник новонародженої дівчинки; Б — зрілий фолікул; В — жовте тіло: Г — яєчник жінки в постменопаузі):
1 — поверхневий епітелій яєчника; 2 — білкова оболонка (tunica albuginea); 3 — яйцеклітина; 4 — ядро; 5 — блискуча зона (zona pellucida); 6 — променистий вінець (corona radiata); 7 — тяжі епітеліальних клітин, що мігрують донизу і оточують яйцеклітину; 8 — яйценосний горбок (cumulus oophorus); 9 — кора яєчника (cortex) ; 10 — гранульозна оболонка фолікула; 11 — внутрішня тека-оболонка фолікула (iht'r.fi interna), 12 — зовнішня тека-оболонка фолікула (theca externa); 13 — при- мордіальний фолікул; 14 — епітеліальний тяж; 15 — первинний фолікул; 16 — порожнина зрілого фолікула, наповнена фолікулярною рідиною (antrum); 17 — біле тіло (corpus albicans); 18 — гілюсні клітини; 19 — кровоносні судини, що входять у ворота яєчника; 20 — поверхневий епітелій; 21 — фолікул, що розвивається; 22 — зрілий фолікул; 23 — геморагічне жовте тіло (розірваний фолікул); 24 — звільнена яйцеклітина; 25 — раннє жовте тіло; 26 — лютеїнові клітини; 27 — фібрин; 28 — кров'яний згусток; 29 — зріле жовте тіло; 30 — біле тіло; 31 — фолікулярна порожнина (зі згустком і фібрином); 32 — фолікул у стадії ранньої атрезії; 33 — пізні ат- ретичні фолікули; 34 — гранульозні лютеїнові клітини; 35 — тека-лютеїнові клітини; 36 —зовнішня тека-оболонка естрадіолу, через 32 год після початку зростання рівня ЛГ і між 12 і 16 год після досягнення піка ЛГ у сироватці крові.
Хвиля ЛГ у середині циклу ініціює завершення метафази першого ділення. Ооцит вступає у метафазу ІІ ділення і утворюється перше полярне тільце. Завершення мейозу і утворення другого полярного тільця відбуваються тоді, коли яйцеклітина пенетрується сперматозоїдом. ЛГ стимулює синтез простагландинів і протеолітичних ензимів. Плазмін під дією ФСГ сприяє процесу виштовхування ооцита і куму- люса під час розриву фолікула. Звільнений ооцит потрапляє в порожнину маткової труби за допомогою рухів її торочок і далі в матку.
Після овуляції кількість фолікулярної рідини, діаметр і об’єм фолікула зменшуються. Фолікулярна стінка скручується, місце розриву фолікула — стигма овуляції — закривається фібрином. Якщо запліднення не відбувається, утворюється жовте тіло менструації (corpus luteum). Під дією ЛГ жовте тіло вступає у стадію розквіту і секретує прогестерон (20 мкг/добу), а також естрадіол. Прогестерон забезпечує фазу секреції в менструальному циклі, сприяє підготовчим змінам у слизовій оболонці матки, спрямованим на здійснення імплантації, забезпечує фізіологічний перебіг вагітності. Високий рівень ЛГ, а також ХГЛ необхідні для підтримки секреторної функції жовтого тіла. Стадія розквіту жовтого тіла триває 12-14 днів; діаметр жовтого тіла менструації досягає 1,5-2 см. Якщо імплантації не відбувається, жовте тіло підлягає зворотному розвитку. Його клітини підлягають жировій дегенерації, і утворюється біле тіло (rnrpus albicans), яке з часом абсорбується стромою яєчника.
Рівень прогестерону рівномірно зростає у сироватці крові після овуляції і досягає плато через 1 тиж і потім зменшується, якщо не виникає імплантації. Зростаючі рівні естрогенів та прогестерону за механізмом негативного зворотного зв’язку пригнічують секрецію ФСГ і ЛГ. Естрадіол інгібує перважно ФСГ (негативний зворотний зв’язок), прогестерон — ЛГ. Продукція прогестерону жовтим тілом спричинює місцевий лютеолітичний ефект. Вважають, що зростання рівня прогестерону в яєчнику запобігає дозріванню фолікула в цьому яєчнику в наступному циклі.
Після закінчення лютеолізу і зменшення рівнів естрадіолу і прогестерону виникає слабший зворотний зв’язок. Перед початком менструації починають зростати рівні ФСГ і ЛГ для подальшої стимуляції росту фолікулів у наступному яєчниковому циклі.Естрадіол і прогестерон чинять пряму пригнічуючу дію на синтез і секрецію гіпофізарних гонадотропінів, а також на вивільнення ГнРГ, змінюючи частоту та амплітуду його імпульсів. Цей стероїдний вплив здійснюється за допомогою нейро- трансмітерів (допаміну та норепінефрину) і нейромодуляторів (в-ендорфінів) в ар- куатних ядрах гіпоталамуса. В ранню фолікулярну фазу пульсація ЛГ відбувається кожні 90 хв, припиняється під час сну і зростає в середній та пізній фолікулярній фазі (1 імпульс за 60 хв вдень і вночі) і уповільнюється до 1 імпульсу за 90 хв у ранню лютеїнову фазу і 1 імпульсу за 3 год у пізню лютеїнову фазу. Подібні зміни пульсації ФСГ у периферичній крові не відбуваються, можливо, у зв’язку з його довшим періодом напівжиття.
Зростання частоти імпульсів ЛГ у пізню фолікулярну фазу є важливим для стимуляції секреції естрадіолу. Близько 80 % пульсуючих викидів ЛГ відбуваються після підйому рівня естрогенів у периферичній крові (позитивний зворотний зв’язок). Пульсація ЛГ в середню і пізню лютеїнову фази стимулює пульсуючу продукцію прогестерону. Зменшення частоти імпульсів ЛГ протягом середньої лютеїнової фази може відбуватися внаслідок негативної зворотної дії прогестерону і естрадіолу. Зменшення частоти секреції ЛГ пов’язують із впливом прогестерону на вивільнення ГнРГ гіпоталамусом. Медіаторами цього процесу є, можливо, в-ендорфіни. В свою чергу зменшення амплітуди імпульсів ЛГ пов’язують із негативною зворотною дією прогестерону на гіпофіз.
Екскреція ФСГ і ЛГ із сечею коливається від 1 до 10 МО/добу, а їх рівень у сироватці крові — від 1 до 100 мМО/мл. Екскреція естрогенів (естрону, естрадіолу і естріолу) є найнижчою протягом ранньої фолікулярної фази, досягає максимуму безпосередньо перед піком ЛГ, після чого швидко знижується і знову підвищується в лютеїнову фазу, наприкінці якої також знижується.
Зростання рівня цих естрогенів у лютеїнову фазу має меншу амплітуду, але більшу тривалість, ніж протягом передову- ляторного піка. Пік екскреції естрогенів із сечею становить 50-75 мкг/добу. Сироватковий рівень естрадіолу зростає з менш ніж 50 пг/мл у ранню фолікулярну фазу до 200-500 пг/мл у середині циклу з підвищенням у лютеїнову фазу до 100-150 пг/мл.Основним метаболітом прогестерону, який екскретується з сечею, є прегнандіол. Рівень прегнандіолу не перевищує 0,9 мкг/добу перед овуляцією і вищий 1 мкг/добу (3-4 мкг/добу) після овуляції. Рівень прогестерону у сироватці крові менший 1 нг/мл перед овуляцією і в середині лютеїнової фази досягає 10-20 нг/мл. За 46 днів до початку менструації рівні естрадіолу, прогестерону і 17-гідроксипрогестеро- ну починають знижуватись. У середині циклу спочатку підвищується рівень естрадіо- лу. Після досягнення піка естрадіолу виникає раптовий підйом хвилі ЛГ і ФСГ. Секреція ЛГ досягає піка через 18 год і утримується (плато) протягом 14 год, а потім знижується. Середня тривалість хвилі ЛГ дорівнює 24 год. За 12 год до початку хвилі ЛГ зростає рівень прогестерону і 17-гідроксипрогестерону. Разом з піком ЛГ спостерігається зниження концентрації естрадіолу і подальше зростання рівня прогестерону. Таке зрушення стероїдогенезу на користь прогестерону замість продукції естрадіолу відбувається внаслідок лютеїнізації гранульозних клітин фолікула під дією ЛГ.
Рівні інших гормонів протягом яєчникового циклу також підлягають деяким змінам. Сироваткові рівні андростендіону і тестостерону незначно коливаються протягом циклу, хоча їх середні рівні є вищими протягом фолікулярної фази порівняно з лютеїно- вою. Рівень ТТГ є відносно стабільним, тимчасом як вміст адренокортикотропного гормону (адренокортикотропіну, АКТГ) і гормону росту мають передовуляторний пік, а рівень пролактину дещо підвищується у лютеїнову фазу циклу.
Якщо відбувається запліднення, продукція прогестерону жовтим тілом триває під стимулювальним впливом ХГЛ, який секретується синцитіотрофобластом і утворюється жовте тіло вагітності. Жовте тіло досягає в діаметрі 1,5-2 см і зазнає зворотного розвитку через 11-12 тиж.