Яєчники
Гонадна регуляція репродуктивної системи факторами росту
Інсуліноподібні фактори росту (IGFs) представлені сімейством з двох пептидів: IGF-І і IGF-ІІ, які мають структурну подібність до проінсуліну і продукуються в різних ділянках тіла.
Їх рівень у крові не змінюється протягом менструального циклу; отже вони діють аутокринним/паракринним механізмами, зв’язуючи рецептори І і ІІ типів до IGF у різних клітинах-мішенях. Рецептори типу І є більш активними. Дію IGFs на органи-мішені регулює IGFs-зв’язуючий протеїн (IGFBPs).Базальний і гонадотропінзалежний стероїдогенез у тека- і гранульозних клітинах стимулює IGF-І. Останній сприяє ФСГ-залежному зростанню рівня цАМФ, ЛГ-ре- цепторів, протеогліканів і базальному синтезу інгібіну в гранульозних клітинах. Первинним місцем продукції IGF-І в яєчниках є гранульозні клітини, хоча специфічні рецептори мають як гранульозні, так і тека-клітини, що підтверджує можливий регуляторний механізм двокомпонентного яєчникового стероїдогенезу. Рецептори IGF- ІІ знаходяться в гранульозних клітинах IGF-ІІ секретується гранульозними клітинами і посилює стероїдогенез. Хоча інсулін не виробляється в яєчниках, він може впливати на яєчниковий стероїдогенез кількома шляхами. Інсулін може взаємодіяти з рецепторами гранульозних клітин і зв’язувати рецептори IGF-І у високих концентраціях. Інсулін може регулювати рівень циркулюючого IGF-І-зв’язуючого протеїну (IGFBPs), який зменшений у жінок із синдромом полікістозних яєчників, інсу- лінорезистентністю і гіперінсулінемією. Також ймовірно, що андрогенні прояви цих синдромів є вторинними у відповідь на дію інсуліну через IGF-І рецептори в стромі яєчників.
Інгібін, активін і фолістатин
Існують докази, що інгібін, активін і фолістатин відіграють важливу роль у яєчниковому стероїдогенезі. Інгібін і активін — глікопротеїни, які складаються з двох субодиниць, з’єднаних між собою дисульфідними зв’язками.
Два біоактивних інгібіни — інгібін А та інгібін В, кожний молекулярною масою 32 000 дальтон, мають подібну біологічну функцію. Вони синтезуються переважно в гранульозних клітинах, яєч- кових Сертолі-клітинах, у жовтому тілі та плаценті. Продукція інгібіну збільшується під дією ФСГ, хоча її можуть порушувати деякі аутокринні та паракринні фактори. Інгібін більше пригнічує виділення ФСГ, ніж ЛГ. Локальна дія інгібіну на рівні яєчників полягає у стимуляції продукції андрогенів тека-клітинами і пригнічуючому впливі на дозрівання ооцитів. Протягом менструального циклу рівень інгібіну в ранню і середню фолікулярну фазу є низьким, він підвищується через кілька днів після піка ЛГ і досягає максимуму в середині лютеїнової фази. Рівень інгібінів різко зменшується в перименопаузі та менопаузі, що може мати зв’язок з підвищенням рівня ФСГ у цьому періоді життя.Біоактивні форми активіну мають молекулярну масу 28 000 дальтон. На рівні гіпофіза активін стимулює виділення ФСГ, на відміну від інгібіну. Паракринна/ аутокринна функція активіну включає ароматазну активність і продукцію прогестерону, сприяння фолікулогену шляхом диференційованої дії на гранульозні клітини і попередження передчасної лютеїнізації фолікула.
Фолістатин кодується одним геном і не має структурної подібності з активіном і інгібіном. Його дія полягає у пригніченні виділення ФСГ шляхом зв’язування акти- віну, і отже, зменшенні активності активіну. Більш того, фолістатин інгібує численні пара- та аутокринні функції активіну і прискорює темп дозрівання ооцитів шляхом впливу на гранульозні клітини.
Трансформуючий фактор росту (TGF-α) та епідермальний фактор росту (EGF- а) є гомологічними в-субодиниці молекул інгібіну й активіну і є потенційними регуляторами проліферації та диференціації гранульозних клітин.
Ітерлейкін-1 (IL-1) — поліпептидний цитокін, який секретується переважно макрофагами. В яєчниках він утворюється тека-інтерстиційними клітинами і гранульозними клітинами після розриву фолікула.
Регуляція IL-1 на рівні яєчника первинно детермінується локальною концентрацією прогестерону. IL-1 проявляє антигона- дотропну активність шляхом супресії функціональної та морфологічної лютеїнізації гранульозних клітин. Існують докази, що IL-1 може відігравати центральну роль у передовуляторній підготовці та розриві фолікула.Простагландини. Попередником біосинтезу простагландинів (ейкозаноїдів) є арахідонова кислота, яка вивільнюється з мембранних фосфоліпідів за допомогою ферменту ліпази. Ейкозаноїди мають широкий спектр біологічних ефектів і відіграють
важливу роль у функції репродуктивної системи та фізіології яєчників. Вони контролюють процес початку росту фолікула шляхом сприяння посиленню кровотоку навколо фолікула і активують ФСГ-рецептори гранульозних клітин преовулятор- них фолікулів. Простагландини Е2 і F2α (PGB2 і PGF2α) концентруються у фолікулярній рідині преовуляторного фолікула і можуть допомагати процесу овуляції шляхом сприяння активності протеолітичних ензимів у стінці фолікула. Простагландини можуть допомагати регулюванню періоду життя жовтого тіла. PGE2 є, можливо, лютеотропним, а PGF2α чинить лютеолізуючу дію.
Простагландини також мають потенціюючий ефект на рухливість маткових труб, підсилюють стимулюючий ефект естрогенів та інгібують ефект прогестерону на кон- трактильну здатність мускулатури маткових труб.
Численні простаноїди синтезуються в ендометрії: PGE2 і PGF2α PGI2 і тромбоксан A2 (ТХА2). Концентрація PGE2 і PGF2α прогресивно зростає від проліфератив- ної до секреторної фази, найвищий рівень їх спостерігається під час менструації. Ці простагландини допомагають регулювати контрактильність міометрія і регулюють процес менструації.
Яєчникові стероїди
Біохімія. Стероїди є ліпідами, що мають основну хімічну структуру — ядро, яке містить 3 шестивуглеводні кільця, приєднані до 5-вуглеводного атому (циклопен- танпергідрофенантрен, або гонан) (див.
рис. 4.3). Молекулярна маса гонана дорівнює 250-550 дальтон. Такі стероїди, як естрадіол і прогестерон нерозчинні у воді, але швидко розчинюються в органічних розчинниках (хлороформ, діетиловий ефір). Навпаки, стероїди, які мають приєднану сульфатну або глюкуронідну групу (кон’ю- говані стероїди), такі як дегідроепіандростерон сульфат (ДГЕАС) і прегнандіол глю- куронід, є водорозчинними.Першим кроком біосинтезу яєчникових стероїдів є трансформація холестеролу до прегнонолону; з прегнонолону під дією ЛГ синтезуються за допомогою специфічних ферментів дегідроепіандростерон, прогестерон, 17-оксипроксипрогестерон, ан- дростендіон і тестостерон (див. рис. 4.4). Андростерон і тестостерон взаємодіють між собою і конвертуються в естрон і, пізніше, в естрадіол за допомогою ферменту ароматази. Цей ферментативний процес ароматизації відбувається внаслідок втрати С-19-метилової групи і утворення ароматичного кільця на С-18.
Яєчник секретує три первинних стероїди: естрадіол, прогестерон і андростенді- он. Ці гормони є основними секреторними продуктами зрілого фолікула, жовтого тіла і строми яєчника відповідно (рис. 4.5, 4.6). Яєчник в деякій кількості секретує також прегненолон, 17-оксипрогестрон, тестостерон, ДГЕАС і естрон. У зв’язку з тим, що яєчники не мають ферментів 21-гідроксилази, 11-гідроксилази і 18-гідрок- силази, вони не можуть синтезувати мінералокортикоїди або глюкокортикоїди. Кожного дня яєчник секретує 0,1-0,5 мг естрадіолу; його кількість зменшується перед менструацією і зростає напередодні овуляції. Щоденна продукція прогестерону варіює від 0,5 мг у фолікулярну фазу до 20 мг у лютеїнову фазу. Протягом фолікулярної фази майже весь прогестерон синтезується наднирковими залозами і дуже мала кількість — яєчниками. Яєчник синтезує 1-2 мг андростендіону, менше ніж 1 мг ДГЕА і близько 0,1 мг тестостерону щоденно. Крім гонадного біосинтезу стероїдів, їх метаболізм здійснюється також в екстрагонадних тканинах (інтерконверсія андро- стендіону і тестостерону, естрону і естрадіолу), переважно за рахунок оксидації стероїдів у їх попередники, які мають меншу біологічну активність.
Естрон також конвертується в естрон сульфат, який має довгий період напівжиття і є найбільшим компонентом пулу циркулюючих естрогенів. Хоча естрон сульфат не є біологічно активним, фермент сульфатази в таких тканинах, як молочна залоза і ендо- метрій, може конвертувати його до естрону, який у подальшому конвертується до естрадіолу.Андростендіон у жировій тканині може конвертуватися в естрон (1,3 % денної кількості), що в більшій мірі притаманний жінкам з ожирінням (до 7 %).
Транспорт. Після виділення в кро- вотік стероїди зв’язуються специфічним протеїном, — глобуліном, що зв’язує статеві гормони (SHBG) і кортикостероїд- зв’язуючим глобуліном (CBG), або з неспецифічним протеїном, наприклад альбуміном. Зв’язані форми стероїдних гормонів становлять близько 95 % усього циркулюючого гормону: решту називають «вільним» гормоном. Наприклад, у жінок у пременопаузальному періоді близько 65 і 30 % циркулюючого тестостерону зв’язані SHBg та альбуміном; лише 2 % є незв’язаними. Альбумін зв’язує всі стероїди, SHBG — переважно зв’язує дигідротестостерон, тестостерон і ес- традіол (за порядком зменшення афін- ності). Зв’язується з більшою афінністю до кортизолу, кортикостерону і, в меншій мірі, прогестерону. Циркулюючі рівні кожного глобуліну зростають під дією естрогенів; рівень SHBG також збільшується при ожирінні та гіпертиреоїдизмі і зменшується під впливом андрогенів і гіпотиреоїдизму.
Метаболізм. Основними місцями метаболізму стероїдів є печінка і, в меншій мірі, нирки. Близько 10-15 % прогестерону трансформується у прегнандіол — 3- глюкуронід, який є основним метаболітом прогестерону в сечі. Естрадіол і естрон конвертуються в печінці до естріолу. Ці три естрогени часто називають класичними естрогенами, тому що вони були виділені першими. Ці естрогени в печінці і слизовій оболонці тонкої кишки кон’югу- ються у різні форми естроген-сульфатів і глюкуронідів (естрон сульфат, естрадіол- 17-глюкуронід, естріол-16 глюкуронід та ін.).

Рис.
4.5. Зміни концентрації гіпофізарних і яєчникових гормонів протягом яєчникового циклу:1 — ЛГ, мМО/мл; 2 — ФСГ, мМО/мл; 3 — П (прогестерон), нг/мл; 4 — Е2(естрадіол), пг/мл; 5 — 17-ОГП (гідроксипрогестерон, нг/мл)
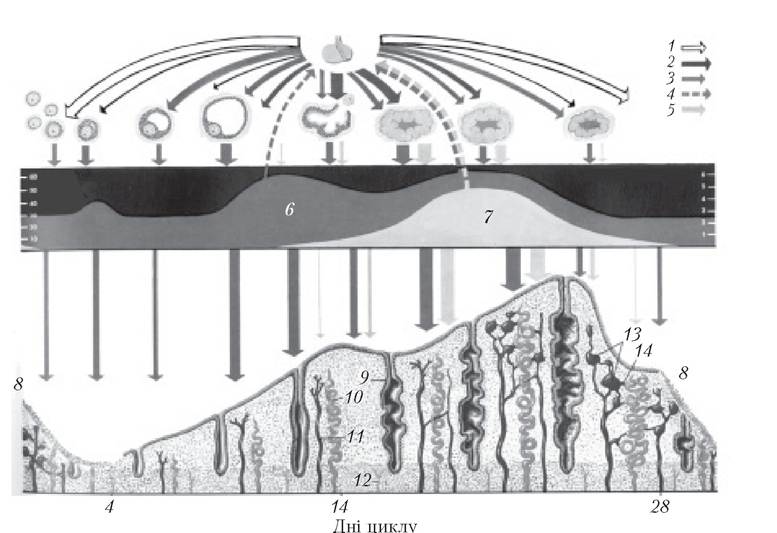
Рис. 4.6. Яєчниковий цикл:
1 — ФСГ; 2 — ЛГ; 3 — естрадіол; 4 — інгібін; 5 — прогестерон; 6 — рівень естрогенів у крові; 7 — рівень прогестерону в крові; 8 — менструація; 9 — залоза; 10 — спіральна артерія; 11 — вена; 12 —базальна артерія; 13 — венозні лакуни; 14 —анастомоз
Дія гормонів. На відміну від мембранних рецепторів протеїнових гормонів, рецептори стероїдних гормонів є внутрішньоклітинними. Рецептори стероїдних гормонів зв’язують специфічні класи стероїдів. Так, естрогенові рецептори зв’язують натуральні та синтетичні естрогени, але не прогестини або андрогени. Афінність рецепторів до стероїдів корелює з активністю останніх. Наприклад, естрогенові рецептори мають більшу афінність до естрадіолу, ніж до естрону та естріолу. Після зв’язування стероїдного гормону (С) зі своїм рецептором (Р) утворюється гормон-рецеп- торний комплекс (СР). Рецептори стероїдних гормонів локалізуються як у цитоплазмі, так і в ядрі клітини. Вони утримуються у неактивному стані шляхом з’єднання з білками тіплового шоку. Зв’язуючись із гормонами, вони вивільнюють білки тіплового шоку і трансформуються так, що можуть зв’язуватися з ядерною ДНК. м-РНК шляхом транскрипції утворюється з сегмента ДНК, мігрує у цитоплазму до рибосом і передає інформацію щодо синтезу нового білка. Сила сигналу для клітини залежить від концентрації гормонів, їх рецепторів, а також ступеня афінності рецепторів до гормонів.
Естрогени стимулюють синтез як естрогенових, так і прогестеронових рецепторів у тканинах-мішенях, таких як ендометрій. Прогестини пригнічують синтез як естрогенових, так і прогестеронових рецепторів. Отже, вміст естрогенових та прогестеро- нових рецепторів у ендометрії досягає піка в середині циклу і потім зменшується. Мітотична активність і швидкість росту ендометрія також є максимальними в середині циклу. Прогестини посилюють внутрішньоклітинний синтез естрадіол-дегідро- генази, яка конвертує більш активний естрадіол до менш активного естрону, що прогресивно зменшує естрогенну активність у клітинах-мішенях.
Антиестрогени, такі як, наприклад, кломіфен або тамоксифен, зв’язують естрогенові рецептори, але ініціюють невелику транскрипцію. Отже, естрогенові рецептори виснажуються без нового синтезу рецепторів або дії естрогенів, що призводить до виснаження естрогенових рецепторів.