Гіпофіз
Структура і функція гонадотропінів
Глікопротеїнами з високою молекулярною масою є ЛГ і ФСГ (28 000 і 37 000 дальтон, відповідно). Вони мають однакову а-субодиницю (14 000 дальтон), яка складається з 90 амінокислот і є подібною до структури а-субодиниці ТТГ і ХГЛ.
в-субодиниці цих гормонів мають різні амінокислоти і карбогідрати і відповідають за специфічну біологічну активність. Субодиниці а і β приєднуються за допомогою сульфгідрильних груп. ЛГ має період напівжиття 30 хв, а ФСГ — 3,9 год. ЛГ діє на тека-клітини фолікула і стимулює синтез ними стероїдів, тимчасом як ФСГ діє переважно на гранульозні клітини і стимулює зростання фолікулів. ФСГ виділяється в більшій кількості, ніж ЛГ до пубертатного періоду. При встановленні нормального менструального циклу секреція ЛГ перевищує виділення ФСГ. Після менопаузи співвідношення ЛГ/ФСГ знову зменшується. Це вибіркове пригнічення виділення ФСГ протягом репродуктивного періоду відбувається під дією зростаючих рівней естрадіолу та інгібіну.Рецептори ЛГ присутні у тека-клітинах протягом усього менструального циклу, у гранульозних клітинах вони з’являються після дозрівання фолікула під впливом ФСГ і естрадіолу; вони присутні у жовтому тілі. Кожна гонадна клітина-мішень має 200030 000 мембранних рецепторів. Максимальна стимуляція гормональної активності відбувається, коли щонайменше 5 % цих рецепторів зв’язані з гормоном. Основною функцією ЛГ є стимуляція синтезу андрогенів тека-клітинами і синтезу прогестерону жовтим тілом шляхом сприяння внутрішньоклітинній продукції цАМФ. Механізм дії ЛГ на гранульозні клітини остаточно не встановлений, але він, можливо, діє синергічно з ФСГ і стимулює дозрівання фолікула. ЛГ стимулює також транспорт амінокислот і синтез РНК в яєчниках. ЛГ може стимулювати овуляцію шляхом індукції активатора плазміногена, який зменшує міцність оболонки фолікула перед його розривом (рис.
4.4).Рецептори ФСГ локалізуються переважно в мембрані гранульозних клітин. Додатково до стимуляції ЛГ-рецепторів на цих мембранах ФСГ активує ароматазу і 3в-гідроксистероїд-дегідрогеназу всередині клітин шляхом активації цАМФ. Стимуляція ФСГ ізольованих гранульозних клітин in vitro приводила до виділення незначної кількості естрогенів, але при додаванні андрогенів або тека-клітин синтез естрогенів значно зростав. Ці дані підтверджують двоклітинну гіпотезу продукції
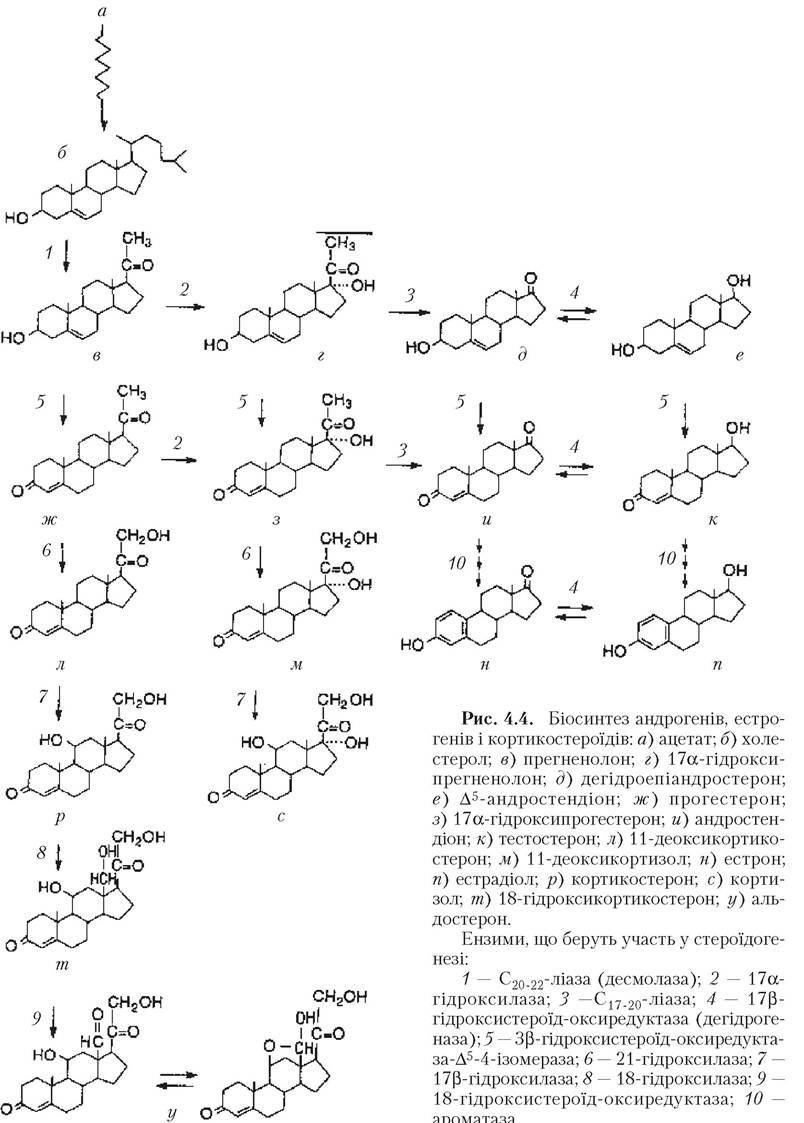
естрогенів. Згідно з цією гіпотезою, ЛГ діє на тека-клітини, що продукують андрогени (андростендіон і тестостерон). Андрогени потім транспортуються в гранульозні клітини, де під дією ФСГ ароматизуються в естрогени (естрон і естрадіол) за допомогою ферменту ароматази.
Під дією естрогенів у гранульозних клітинах стимулюються мітози; кількість клітин збільшується. Зростає кількість рецепторів естрадіолу та ФСГ, що підтримується внутрішньоклітинним рівнем цАМФ навіть при зменшенні рівня ФСГ у крові. В гранульозних клітинах ЛГ діє синергічно з ФСГ щодо зростання ЛГ-рецеп- торів та ініціації лютеїнізації фолікула, що призводить до зростання рівня прогестерону. Передчасне вивільнення ЛГ може перервати цей процес і призвести до передчасної лютеїнізації фолікула. Здатність фолікула відповідати на дію естрогенів визначає, досягне він зрілості чи відбудеться його атрезія.
Синтез простагландинів шляхом внутрішньоклітинної продукції цАМФ стимулює також ЛГ. Простагландини можуть відігравати роль у розриві фолікула. Вміст простагландинів у фолікулі зростає під час підвищення рівня гонадотропінів і може призводити до стимуляції скорочення гладких м’язів. Прогестерон посилює активність протеолітичних ферментів, які діють разом з простагландинами у напрямку деградації та розриву фолікула. Концентрація активатора плазміногена (АП) під дією ЛГ зростає у передовуляторному фолікулі. Призначення інгібіторів АП блокує овуляцію.
На рівні яєчників вибір і початкове зростання фолікулів спостерігається незалежно від гонадотропних гормонів. Хоча кількасот фолікулів, можливо, починають зростати, більшість із них дегенерує. Не більше 30 фолікулів стають гонадотропінзалеж- ними і присутні на початку менструального циклу. З них за фізіологічних умов лише декілька, з оптимальною ФСГ/ЛГ стимуляцією, відбираються для подальшого росту і розвитку. Вважають, що збереження фолікула від дегенерації під дією ФСГ досягається шляхом редукції андрогенного впливу і підтримки естрогензалежного розвитку. Спочатку це відбувається шляхом посилення продукції активіну, а потім шляхом метаболізації ЛГ-індукованих текальних андрогенів у гранульозних клітинах. Вважається, що вибір домінантного фолікула визначається його більшою чутливістю до ФСГ і здатністю до продукції значної кількості естрогенів, а також можливістю модулювати секрецію гонадотропінів. Домінантний фолікул зазвичай визначається на 7-й день циклу.