Ингибирование активности липолитических ферментов хитозаном в гетерогенных растворах, на границе раздела фаз
Изучение ингибирования низкомолекулярным хитозаном липолитической активности было проведено на трех липазах - свиной панкреатической липазе, липазе из С. rugosa и липазе из проростков пшеницы в гетерогенной фазе, на границе масло-вода.
Зависимости начальных скоростей гидролиза оливкового масла липазами от концентрации субстрата в отсутствие и в присутствие ингибитора описываются уравнением Михаэлиса-Ментен, после линеаризации, которого в координатах Лайнуивера-Берка были определены максимальные скорости гидролиза и константы Михаэлиса Кт. На рис. 16 показана зависимость обратной начальной скорости гидролиза оливкового
масла свиной панкреатической липазой от концентрации субстрата в отсутствие и в присутствие хитозана. Аналогичные зависимости получены и для липазы из проростков пшеницы и для липазы из С. rugosa.
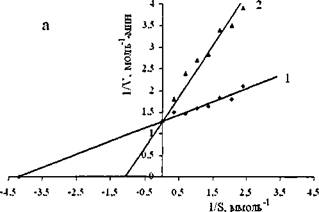
Рис. 16. График зависимостей в обратных координатах начальных скоростей гидролиза оливкового масла свиной панкреатической липазой от концентрации субстрата в отсутствии (1) и в присутствии ингибитора (2) при pH 6,7 и ЗГС.
Проекция прямых на ось абсцисс позволила определить, что хитозан является конкурентным ингибитором липолитических ферментов. Найденные из этих зависимостей значения молекулярных констант скорости, kcat и констант Михаэлиса, Кт представлены в таблице 20.
Таблица 20.
Кинетические константы реакции гидролиза оливкового масла липолитическими ферментами
| Фермент | Кт, моль | K^atjCek | Kcat/Km, моль’-сек'1 |
| Панкреатическая липаза свиная | 0,24*10'3 | 19,4 | 81*103 |
| Candida rugosa | 0,27*10'3 | П,4 | 42,2*103 |
| Из проростков пшеницы | 0,2*10’3 | 1,7 | 8,5 • 103 |
Эффективность гидролазного действия панкреатической липазы на оливковое масло, выражаемая бимолекулярной константой kcat/Km, в 10 раз
выше, чем для растительной липазы, и в 5 раз выше, чем для липазы из С.
rugosa. Разница в эффективности гидролазного действия на субстрат между панкреатической липазой и липазой из С. rugosa составляет около двух раз. Это обусловлено различиями в величинах как kcat, так и Км.Из данных табл. 21 следует, что низкомолекулярный хитозан ингибирует липолитическую активность, константы ингибирования Kj изменяются в пределах 70-160 мкМ, в зависимости от ингибируемого фермента.
Таблица 21.
Константы ингибирования липолитических ферментов
низкомолекулярными хитозанами и моноглюкозамином
| Фермент | Константы ингибирования Kj, моль | ||
| Хитозан | N-сукцинил хитозан | Моноглюкозамин | |
| Панкреатическая липаза свиная | 7,7*10'5 | 1,1*10"5 | 4,3*10’2 |
| Candida rugosa | 1,52’КГ4 | 3,3*10’4 | 1,1*10'2 |
| Из проростков пшеницы | 1,14*10"4 | 4,2*10‘4 | 0,36*10'2 |
Значения констант ингибирования растительной липазы и липазы из С. rugosa одного порядка. Константа ингибирования хитозаном панкреатической липазы на порядок ниже и составила 77 мкМ. Увеличение констант ингибирования хитозаном растительной и грибной липаз по сравнению с константами, найденными при гидролизе водорастворимого субстрата, согласуется с данными о том, что при нахождении липолитических ферментов в зоне раздела фаз, липид - вода, эффективность связывания с ингибитором существенно увеличивается за счет значительного преобладания открытой конформации фермента.
Ингибирование активности липолитических ферментов моноглюкозамином
В качестве ингибитора липолитической активности был изучен и моноглюкозамин.
Ранее было показано, что моноглюкозамин не ингибируетлиполитические ферменты [243]. Представление полученных нами экспериментальных данных в координатах Лайнуивера-Берка (рис. 17.) позволило определить константы ингибирования изучаемых липаз.
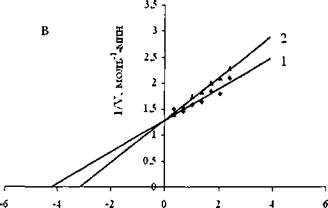
1/S, ммоль'1
Рис.17. График зависимостей в обратных координатах начальных скоростей гидролиза оливкового масла свиной панкреатической липазой от концентрации субстрата в отсутствии (1) и в присутствии (2) моноглюкозамина при pH 6,7 и 37°С.
Эффективность ингибирующего действия моноглюкозамина оказалась на три порядка ниже, в случае ингибирования панкреатической липазы и на два порядка ниже, в случае ингибирования растительной липазы и грибной липазы. Константы ингибирования, К; составили 10-40 мМ (табл.21). Увеличение значений констант ингибирования связано, по-видимому, с участием в комплексообразовании с липазами только одного единственного звена моноглюкозамина. Данный вывод подтверждают найденные значения свободной энергии, AG комплексообразования моноглюкозамина с липазами.
Вклад свободной энергии, AG в комплексообразование между липазами и моноглюкозамином составил -2,32 ккал/моль для панкреатической липазы, -2,41 ккал/моль для липазы из проростков пшеницы и -2,7 ккал/моль для липазы из С. rugosa. Константы ингибирования липаз низкомолекулярным хитозаном со свободными аминогруппами составляют от 70 - 160 мкмоль, что соответствует значениям AG -5,8 ккал/моль для свиной панкреатической
липазы, -5,0 ккал/моль для липазы из проростков пшеницы и -5,4 ккал/моль для липазы из С. rugosa. Разница в значениях свободной энергии комплексообразования с липазами моноглюкозамина и хитозана отличается в два раза, что вероятнее всего свидетельствует о том, что при гидрофобном взаимодействии происходит связывание как минимум двух звеньев хитозана с липолитическими ферментами, а в случае взаимодействия моноглюкозамина, связывается одно единственное звено.
Таким образом, мы показали, что низкомолекулярный хитозан подавляет активность липолитических ферментов, как в гомогенных растворах, так и на границе раздела фаз. Низкомолекулярный хитозан является конкурентным ингибитором свиной панкреатической липазы, липазы из C.rugosa и липазы из проростков пшеницы. Хитозан ингибирует специфическую активность липазы C.rugosa и проростков пшеницы в гомогенных водных растворах со значениями К; порядка 1 мМ. Константы ингибирования, Kj свиной панкреатической липазы, липазы из C.rugosa и липазы из проростков пшеницы на границе раздела фаз масло-вода, составили 77 мкМ, 152 мкМ и 114 мкМ, соответственно. При нахождении липолитических ферментов в зоне раздела фаз, липид - вода, эффективность связывания ингибитора с ферментом существенно увеличивается за счет значительного преобладания открытой конформации липаз. Величина эффекта ингибирования зависела от времени преинкубации хитозана с липазами и достигала максимума при времени преинкубации 30 мин. Хотя хитозан является поликатионом, вклад электростатических взаимодействий в его комплексообразование с липазами незначителен.
Константы ингибирования многих известных ингибиторов липолитических ферментов, того же порядка, что и найденные константы ингибирования хитозаном [238-239]. Однако большинство ингибиторов применяемых для лечения избыточного веса и сопутствующих ему заболеваний обладают рядом побочных эффектов [249-250].
Хитозан благодаря наличию положительного заряда и своему строению
является полифункциональным соединением, обладающим целым рядом уникальных свойств: высокой совместимостью с животными тканями, биодеградируемостью, низкой токсичностью и др. [15-21,25]. Это открывает большие перспективы использования низкомолекулярного хитозана в качестве ингибитора липолитических ферментов для профилактики и лечения нарушений липидного метаболизма и сопутствующих ему заболеваний в составе биологически активных добавок.
выводы
1. Разработана технологическая схема переработки кутикулы личинок восковой моли Galleria mellonella с получением хитин-меланинового комплекса и хитозана различной молекулярной массы.
Определены оптимальные условия для каждой стадии схемы переработки.2. Показано, что хитозан обладает противотуберкулезной активностью. Низкомолекулярные образцы хитозана проявляют максимальную антибактериальную активность. С увеличением молекулярной массы противотуберкулезный эффект хитозана снижается. Установлено, что достаточно 10 минут взаимодействия клеток микобактерий с хитозаном, чтобы наблюдался антибактериальный эффект. Введение отрицательно заряженных групп в молекулу хитозана ведет к утрате противотуберкулезных свойств.
3. Проведено последовательное фракционирование экстракта выделенного из восковой моли. После двух хроматографических стадий очистки - гидрофобной и металл-хелат аффинной удельная липолитическая активность возрастает в 29 раз. С увеличением липолитической активности в результате последовательного фракционирования экстракта возрастает и антибактериальная активность, в отношении атипичного штамма М. smegmatis.
4. Установлено, что хитозан является эффективным конкурентным ингибитором липолитических ферментов, как в гомогенных водных растворах, так и на границе раздела фаз. Кинетические константы ингибирования липаз хитозаном составили 70-160 мкМ.
Еще по теме Ингибирование активности липолитических ферментов хитозаном в гетерогенных растворах, на границе раздела фаз:
- СОДЕРЖАНИЕ
- Ингибирование активности липолитических ферментов хитозаном в гетерогенных растворах, на границе раздела фаз