Ингибирование активности липолитических ферментов хитозаном в гомогенных водных растворах
Для выяснения механизма каталитического действия липолитических ферментов первоначально лучше проводить исследования с водорастворимыми субстратами. При кинетическом анализе липолиза в гомогенной водной фазе не возникает осложнений, которые связаны с наличием поверхности раздела фаз.
Скорости гидролиза водорастворимых субстратов гораздо меньше, чем скорости гидролиза нерастворимых в воде, но достаточно велики, чтобы их можно было определить экспериментально. Нами было изучено взаимодействие низкомолекулярного хитозана с липазой из Candida rugosa и липазой из проростков пшеницы в водных растворах, используя паранитрофенил пальмитат в качестве субстрата.Подбор оптимальных параметров ферментативной реакции в гомогенных водных растворах
В результате изучения влияния Мц хитозана от 5-94 кДа на степень ингибирования активности липазы из С.rugosa, было показано, что максимальное снижение активности наблюдалось при действии хитозана с молекулярной массой 6 кДа и менее (таблица 18). В дальнейших исследованиях мы использовали хитозан с Мц порядка 5-6 кДА. Полученные результаты согласуются с литературными данными, полученными Tsujita Т., о том, что хитозан с Мц 10 кДа ингибирует активность панкреатической липазы лучше, чем хитозан с Мц 46 кДа [243].
Таблица 18.
Зависимость значений констант ингибирования липазы C.rugosa от молекулярной массы ингибитора
| Мц хитозана, кДа | К;,МОЛЬ |
| 94 | 1,4 |
| 56 | 0,5 |
| 23 | 2,5*10 "2 |
| 15 | 8*10 "3 |
| 6 | 1,2*10 3 |
Для оптимизации условий измерения констант ингибирования липаз хитозаном первоначально оценивали оптимальное время преинкубации фермента и ингибитора перед началом ферментативной реакции.
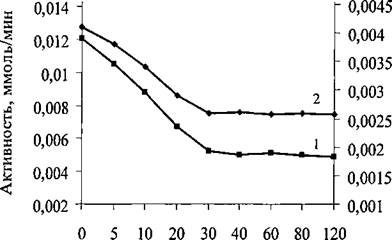
Известно, что в растворах молекула липазы C.rugosa имеет две конформации [244]. В одной из них активный центр фермента закрыт пептидным фрагментом молекулы, а в другой - открыт, что делает его доступным для связывания субстрата и других лигандов. В водных растворах липаза находится преимущественно в неактивной конформации с закрытым активным центром. При нахождении липазы в зоне раздела липид - вода ее активность резко возрастает за счет изменения равновесия между двумя конформациями в сторону значительного преобладания открытой конформации. Предполагается, что ингибиторы липазы C.rugosa взаимодействуют преимущественно, если не исключительно, с открытой формой фермента и что любые переходы между этими двумя конформациями фермента в растворе являются медленными [245]. Поэтому время установления концентрационного равновесия в растворе липазы после добавления ингибитора будет определяться характеристическим временем перехода между закрытой и открытой конформациями фермента. На рис. 11 представлены зависимости активности липазы C.rugosa и проростков пшеницы от времени преинкубации с хитозаном до добавления в реакционную смесь субстрата.Время преинкубации варьировали от 5 до 120 мин. Степень
ингибирования обеих липаз зависела от времени преинкубации вплоть до 30 мин, после чего оставалась примерно постоянной. Поэтому далее величина времени преинкубации соответствовала 30 мин.
Время преинкубации, мин
Рис.11. Зависимость ферментативной активности липазы C.rugosa (1) и проростков пшеницы (2) от времени преинкубации с хитозаном при pH 6,7 иЗО°С
На рис. 12 показана зависимость активности липазы C.rugosa и проростков пшеницы от величины pH.
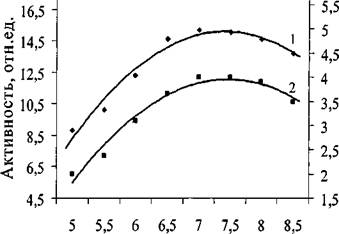
Рис.12. Зависимость ферментативной активности липазы C.rugosa (1) и проростков пшеницы (2) от pH реакционной смеси при 30°С
рн
Максимальная активность ферментов при использовании в качестве
субстрата паранитрофенил пальмитата достигалась при pH 6,7.
Такое же значение pH - оптимума было найдено ранее для липазы из С.rugosa при изучении кинетики гидролиза в водных растворах оливкового масла [246].Последующие измерения константы ингибирования обеих липаз были выполнены при pH 6,7.
Определение кинетических констант
Зависимости начальных скоростей гидролиза паранитрофенил пальмитата липазами от концентрации субстрата в отсутствие и в присутствие ингибитора описываются уравнением Михаэлиса-Ментен, после линеаризации, которого в координатах Лайнуивера-Берка были определены максимальные скорости гидролиза субстрата и константы Михаэлиса Кт.
На рис. 13 приведена зависимость в обратных координатах начальных скоростей гидролиза паранитрофенил пальмитата липазой С.rugosa и проростков пшеницы от концентрации субстрата в отсутствии (прямые 1) и в присутствии хитозана (прямые 2).
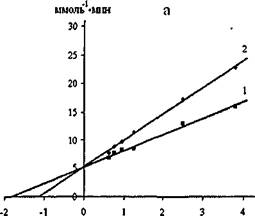
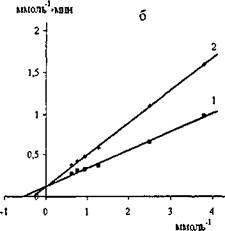
Рис.13. Графики зависимостей в обратных координатах начальных скоростей гидролиза паранитрофенил пальмитата липазой С.rugosa (а) и проростков пшеницы (б) от концентрации субстрата в отсутствии (1) и в
присутствии хитозана (2) при pH 6,7 и 30°С.
Проекция прямых на ось абсцисс позволила определить конкурентный тип ингибирования липаз хитозаном. Найденные значения константы
скорости kcaf, константы Михаэлиса Км и константы ингибирования К,- липаз хитозаном представлены в таблице 19.
Таблица 19.
Кинетические константы реакции гидролиза паранитрофенил пальмитата липазой C.rugosa и проростков пшеницы и константы ингибирования липаз хитозаном
| Источник липазы | Kcat, сек'1 | Км, моль | Kcat/KM( моль'1-сек’1 | Kj, моль |
| Candida rugosa | 2,0 | 64 О'4 | 3,3403 | 1,44 О'3 |
| Зародыши пшеницы | 0,5 | 540"3 | 1,0402 | 0,94 О'3 |
Эффективность гидролазного действия растительной липазы на паранитрофенил пальмитат, выражаемая бимолекулярной константой kcat/KM, в 33 раза ниже, чем для липазы С.
rugosa. Это обусловлено различиями в величинах как kcat, так и Км. Величина К; грибной и растительной липаз хитозаном близки между собой и равны 1,44 О'3 М и 0,94 0‘3 М, соответственно. Такая величина К; типична для большинства известных ингибиторов. Kj панкреатической липазы человека платикодином составляет 0,240'3 М [239], а панкреатической липазы свиньи пропранололом - 0,64О'3 М [247]. При нахождении липазы в зоне раздела липид - вода в эмульсиях эффективность связывания ингибитора с ферментом существенно увеличивалась за счет значительного преобладания открытой конформации фермента [248].С целью выяснения вклада электростатических взаимодействий положительно заряженных групп хитозана с отрицательно заряженными группами белка при образовании комплексов были изучены зависимости констант ингибирования липолитической активности хитозаном от концентрации добавленного в раствор хлорида натрия (рис. 14).
Был проведен расчет числа аминогрупп хитозана, образующих ионную пару с отрицательно заряженной группой белка.
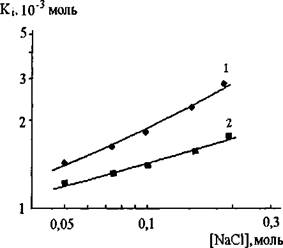
Рис. 14. Зависимость в логарифмических координатах величин Kj липазы C.rugosa (1) и проростков пшеницы (2) хитозаном от концентрации хлорида натрия.
Величина п, численно равная тангенсу угла наклона прямых 1 или 2 к оси абсцисс на рис.14, имела значение 0,44 для липазы C.rugosa и 0,28 для липазы проростков пшеницы. Низкие значения величины п для обеих липаз свидетельствует о незначительном вкладе в свободную энергию комплексообразования электростатических взаимодействий зарядов хитозана и белка. С целью дополнительного подтверждения этого заключения мы провели измерения Kj грибной и растительной липаз низкомолекулярным N-сукцинил хитозаном.
Ингибирование активности липаз низкомолекулярным N-сукцинил хитозаном
В этом производном хитозана каждая сукцинильная группа несет отрицательный заряд.
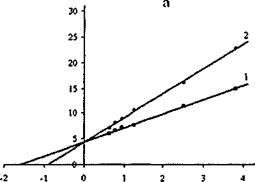
Результаты измерений представлены на рис. 15. Рассчитанные из этих данных К; активностей липазы C.rugosa и проростков пшеницы N-сукцинил хитозаном найдены равными 2,0-10'3 М и 1,7-10'3 М, соответственно.
ммоль1» юш
*1
ммоль «каїн
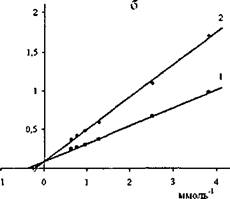
Рис.15. Графики зависимостей в обратных координатах начальных скоростей гидролиза паранитрофенил пальмитата липазой С.rugosa (а) и проростков пшеницы (б) от концентрации субстрата в отсутствии (1) и в
присутствии (2) N-сукцинил хитозана при pH 6,7 и 30°С
Близость значений К; обеих липаз хитозаном со свободными аминогруппами и N-сукцинил хитозаном подтверждает вывод о незначительной роли электростатических взаимодействий при комплексообразовании хитозана с липазами, подавляющего их ферментативную активность.
3.4.2.
Еще по теме Ингибирование активности липолитических ферментов хитозаном в гомогенных водных растворах:
- СОДЕРЖАНИЕ
- Ингибирование активности липолитических ферментов хитозаном в гомогенных водных растворах
- Ингибирование активности липолитических ферментов хитозаном в гетерогенных растворах, на границе раздела фаз