Принцип лімітуючих факторів
Екологічні фактори можуть по-різному впливати на живі організми:
- як подразники вони зумовлюють пристосувальні зміни функцій організму;
- як обмежувальні унеможливлюють існування організмів за певних умов;
- як сигнали вказують на зміни інших факторів середовища.
У впливі факторів середовища на організми та реакціях останніх на цей вплив виявлені певні закономірності, причому вони стосуються факторів будь-якого походження. Кожний екологіч-ний фактор впливає на живі організми позитивно чи негативно, залежно від сили прояву його дії. Сприятливу силу впливу фак-тору, тобто таку, що забезпечує найкращі або оптимальні умови життєдіяльності особин, називають зоною оптимуму еколо-гічного фактора. Будь- яке відхилення від оптимуму негативно впливає на розвиток організмів - чим більші відхилення, тим сильніше пригнічується організм.
Закон оптимуму - фактори позитивно впливають на організми лише в певних межах. Недостатня або надмірна їх дія позначається на організмах негативно (рис. 5).
Зона оптимуму і критичні межі витривалості живих орга-нізмів стосовно якогось екологічного фактора можуть змі-нюватись залежно від того, з якою силою і в якому поєднанні діють одночасно інші екологічні фактори, наприклад, низька
температура і вологість. Цей принцип одержав назву закону взаємодії факторів. Взаємодія екологічних факторів виявля-ється у їх частковій взаємозамінності, наприклад, в'яненню рослин можна запобігти збільшенням вологості ґрунту і зни-женням температури повітря. Проте, взаємна компенсація дії факторів не може бути безмежною і повністю замінити один фактор на інший неможливо.
Якщо хоча б один із екологічних факторів наближається до критичної межі або перевищує її, то, незважаючи на оптимальну дію інших умов середовища, організму загрожує загибель, а цей фактор стає для нього обмежувальним або лімітуючим. Вони можуть бути як абіотичними (наприклад, нестача тепла, вологи або їх надлишок, різка зміна освітленості тощо), так і біотич-ними факторами (наприклад, зайнятість території більш силь-ним конкурентом, нестача запилювачів квітів тощо).
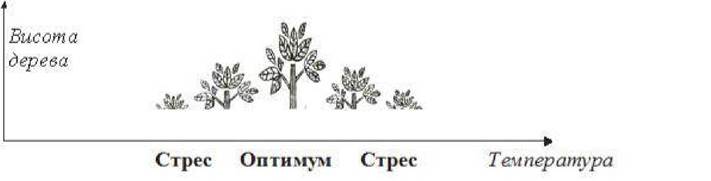
Рис. 5. Оптимальні умови життєдіяльності організмів
Гіпотеза про те, що витривалість організму визначається слабкою ланкою в ланцюзі його екологічних потреб, вперше була висунута і доведена Ю. Лібіхом у 1840 році. Вивчаючи вплив вмісту мікроелементів у ґрунті на ріст рослин він зазначив, що ріст залежить від того елементу, який є в мінімальній кількості. Цей висновок відомий в екології як закон мінімуму. Значення цього закону полягає в тому, що він дає можливість визначити, з чого починати пошук у раз і потреби аналізу умов довкілля.
З часу встановлення Ю. Лібіхом цього закону вченими було відмічено, що сам закон при застосуванні на практиці потребує уточнення. Ю. Одум (1975), для застосування закону мінімуму пропонує користуватись двома допоміжними принципами.
Перший допоміжний принцип: закон Лібіха можна застосовувати без уточнень тільки до умов стаціонарного стану, тобто тоді, коли приток енергії та речовин регулюється її витоком, тобто система знаходиться у стані рівноваги. Перший допоміж-ний принцип Ю. Одума є і обмежувальним. Акцентуючи увагу на ньому, Ю. Одум звертає увагу на те, що система характеризу-ється динамікою і тому, на його думку, введення цього принципу обмежить похибки, які виникають при довготривалих дослі-дженнях екосистем.
Другий допоміжний принцип - торкається взаємодії факторів. Було відмічено, що в певних умовах висока концентрація або доступність певної речовини, або дія другого, не лімітуючого фактору, може змінювати потребу у мінімальній кількості речовини. Прикладом може бути заміна використання молюс-ками кальцію стронцієм, або такий приклад: якщо рослини живуть в умовах малої освітленості, то вони споживають менше Цинку, тощо.
Другий допоміжний принцип, уведений Ю. Одумом, вказує на неможливість аналізу стану системи на основі невеликої кількості елементів. Саме другий допоміжний принцип показує на необхідність комплексного аналізу при будь-якому екологіч-ному дослідженні.
Отже, Ю. Одум відзначив два основних обмеження щодо використання закону мінімуму:
- обмежувальний, тобто закон виконується тільки в статич-них умовах середовища;
- взаємодії факторів, тобто при зміні інших екологічних факторів величина сили дії обмежувального фактора може змі-нюватись.
ЦІКАВО
Закон мінімуму Ю. Лібіха лежить в основі всіх виявлених окремих закономірностей у взаємовідносинах типу „орга-нізм - середовище”. В найпростішому вигляді основний закон мінімуму Ю. Лібіха стосується успішності росту і вели-чини врожаю сільськогосподарської рослини, що залежать від речовини, яка міститься в ґрунті в мінімумі в порівнянні з іншими агрохімічно необхідними речовинами. Цей закон було сформульовано в 1840 р. задовго до виникнення еко-логії як науки. Згодом закон мінімуму було витлумачено як дію будь-якого екологічного фактору, що знаходиться в міні-мумі у порівнянні з іншими екологічними впливами. Іноді закон мінімуму розширюють до правила, що вказує на роль екологічних факторів в розповсюдженні та кількісному розвитку організмів. Це зовнішні умови. Але можлива трактовка закону мінімуму також і „зі сторони” організму: витривалість організму визначається найслабшою ланкою в ланцюгу його екологічних потреб - життєві можливості лімітуються екологічними факторами, кількість та якість яких наближена до необхідного організмові мінімуму. Подальше зниження або погіршення цих факторів веде організм до загибелі. Це „крайня” трактовка закону мінімуму, яка доволі далеко відхиляється від первинного його розуміння, але така, що має більш широке екологічне значення.
Додаткове правило взаємодії факторів в законі мінімуму: організм в певній мірі здатний замінити дефіцитну речовину або інший діючий фактор життя функціонально близькою речовиною чи фактором (наприклад, одну речовину іншою, хімічно близькою) - викликало потік аналогічних постулатів. Серед них закон відносності дії лімітуючих факторів, або закон Лундегарда - Полетаева: форма кривої росту чисельності популяції (її біомаси) залежить не тільки від однієї речовини з мінімальною концентрацією, а й від концентрації та властивостей інших іонів, що є в середовищі.
Вони як лімітуючі фактори складають „системи Лібіха” або „L-систему” (закон сформульовано Лундегардом у 1957 р.).Важливою поправкою та доповненням служить закон неоднозначної (селективної) дії фактора на різні функ-ції організму: будь-який екологічний фактор неоднаково впливає на функції організму, оптимум для одних процесів, наприклад, дихання, не являється оптимумом для інших, наприклад, травлення, і навпаки.
Додаткове правило заміщення дефіцитних речовин закона мінімуму Ю. Лібіха було поглиблене Е. Рюбелем у 1930 р. у вигляді закону (ефекта) компенсації (взаємозамінно-сті) факторів: відсутність або недостатність деяких еко-логічних факторів може бути компенсована іншим близьким (аналогічним) фактором. Наприклад, недостача світла може бути компенсована для рослини достатком вуглекислого газу. Окремим випадком закону відносності дії лімітую-чих факторів Лундегарда - Полетаева та закону компенса-ції факторів Рюбеля е ефект Лундегарда - Стиберга - Якобсона: збільшення біомаси рослини при постійному або навіть зниженому вмісті одного з елементів живлення.
В.В. Альохін сформулював близьке до вищевказаних правило заміщення екологічних умов: будь-яка умова зовнішнього середовища в деякій мірі може заміщуватися іншою; внутрішні причини екологічних явищ при анало- гічому зовнішньому проявленні можуть бути різними.
Наприклад, макрокліматичні впливи можуть бути компенсовані біокліматичними - вічнозелені види ростуть під захистом верхніх ярусів рослинності, багато видів тварин знаходять мікроумови для існування. Це не агрохімічне і гідробіологічне, а широкоекологічне і навіть біогеогра-фічне тлумачення додаткового правила до закону мінімуму Юстуса Лібіха.
Прагненню звузити сферу дії закону мінімуму протисто-їть закон незамінності фундаментальних факторів, запропонований В. Р. Вільямсом. У 1949 р. він сформулював жорсткі обмеження: повна відсутність в середовищі фундаментальних екологічних (фізіологічних) факторів (світла, води, елементів живлення тощо) не може бути замінена іншими факторами.
Цілком очевидно, що будь-який з екологічних компонентів до кінця не може бути замінений - при відсутності енергії нема життя, те саме при повній відсутності води і тому подібне. Для елементарних потреб це очевидно, але в більш складних ситуаціях, особливо коли мова йде про фундаментальні, але не досить відомі та усвідомлені фактори, наприклад, в системі потреб людини, така ймовірність зникає. Це приводить до помилок, іноді фатальних.Закон толерантності. Як показали дослідження Лібіха, існування живого організму зумовлено не тільки недостатністю того чи іншого фактору, але також і їх надлишком. Таким чином, кожен організм має свої межі, які знаходяться між мінімумом та максимумом, тобто оптимум, який забезпечує існування орга-нізму.
Наглядно вплив оптимальних умов на ріст, розмноження та існування організму бачимо на рис. 6. Для кожного виду існу-ють свої межі. Закон про лімітуючу роль максимуму та міні-муму та необхідність оптимальних умов для існування виду ввів В. Шелфорд2з (1913 р.): природним обмежуючим чинником існу-вання організму може бути як мінімальний, так і максималь-ний екологічний вплив, діапазон між якими визначає величину витривалості (толерантності) організму до даного чинника.
В основу екологічної характеристики організмів покладено їх реакцію на вплив факторів середовища: як дуже високі (максимальні), так і дуже низькі (мінімальні) значення факторів середовища можуть бути згубними для організму. Порогове значення даного фактора, вираженого в цифрах, вище або нижче якого організм не може існувати називають критичною точкою.
23 Шелфорд Віктор Ернест (Shelford Victor Ernest) (1877-1968) - американський зоолог, еко-лог, автор найвідомішого у факторіальній екології закону толерантності (1913). Перший прези-дент Екологічного товариства Америки. В 1904- 1914 pp. працював у Чикагському університеті, з 1914 - в Іллінойському університеті (з 1927 - професором). Головні праці в галузі біоценології, теорії та термінології, присвячені методиці польових та лабораторних досліджень.
Крім гідро-біологічних досліджень, вивчав взаємодію організмів у наземних угрупованнях, вплив клі-мату на угруповання, сукцесії; займався класифікацією змішаних угруповань. Першим описав природу Північної Америки з екологічної точки зору.Між цими критичними значеннями і розташована зона еколо-гічної толерантності.
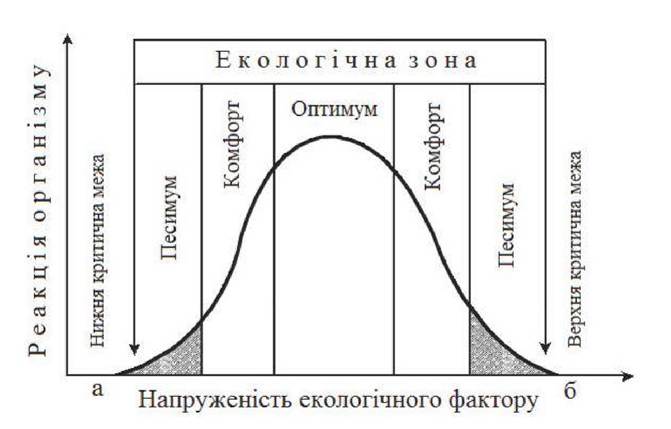
Рис. 6. Критичні межі екологічного фактора:
а - нижня кардинальна точка (екологічний мінімум)
б - верхня кардинальна точка (екологічний максимум)
Організм здатний вижити лише в діапазоні мінливості даного фактора, який ще називають амплітудою. Здатність організму витримувати певну амплітуду коливання фактору називають екологічною валентністю, але для життя організмів значення має не тільки абсолютна величина сили дії фактора, а й швидкість її зміни.
За екологічною валентністю організми поділяють на еврибі-онтних з широкими пристосувальними можливостями, напри-клад, сірий пацюк, горобець, кімнатна муха, таргани тощо, і сте-нобіонтних, які можуть існувати лише у відносно сталих умовах, наприклад, журавель степовий, качкодзьоб тощо. Реакція організму і його адаптаційні можливості до дії фактора, що є
основним чинником еволюції видів, залежить від поєднання дії різних факторів, а для нормального існування організму необхід-ний певний їх набір.
ЗВЕРНІТЬ УВАГУ
Схема стосунків в діапазоні екологічної толерантності була запропонована в 1924 р. німецьким екологом і зоогеогра-фом Р. Гессе, який назвав її валентністю екологічних факто-рів. Варто зазначити, що крива, яка представляє екологічну валентність у межах зони толерантності, не завжди має симетричний вигляд із оптимальною зоною, розташованою в центрі. Наприклад, для прісноводних організмів оптимум знаходиться в нижній межі вмісту солі у воді, тоді як у морських організмів - на протилежному кінці мінливості фактора - в зоні толерантності, де вміст солі найвищий.
Ю. Одум (1975) вводить ряд доповнень до закону толерантності Шелфорда, які стосуються неоднорідності впливу еколо-гічних факторів та реакції на них живих організмів:
- організми можуть мати широкий діапазон толерантності до одного фактору і вузький до іншого;
- організми із широким діапазоном толерантності, як пра-вило, займають панівне становище в біосфері;
- якщо умови існування визначені одним екологічним фак-тором змінюються за межі оптимуму, то змінюються діапазон толерантності до інших екологічних факторів;
- в природі організми часто попадають в умови далекі від оптимально встановлених у лабораторних експериментах;
- період розмноження, росту, як правило є критичним, межі толерантності організму в цей час набагато вужчі, ніж у дорослої особини.
Роз'яснення, наведені Ю. Одумом, багато в чому допомагають при з'ясуванні причин неоднорідності отриманих результатів під час проведення екологічних досліджень. Таким чином, при будь-якому екологічному дослідженні є необхідність детального аналізу не тільки фізико-хімічних умов середовища або ступеню впливу живих організмів одне на одного, але і фаз існування організму.
Для нормального існування організмів їм потрібен комплекс факторів і певний їх режим з допустимими коливаннями вели-чини дії в межах витривалості організмів або толерантності. Організми намагаються жити в певних межах близьких до опти-муму і не досягати критичних меж фактора (рис. 6). У таких умо-вах будь-які організми можуть підтримувати свою чисельність.
Для характеристики амплітуди толерантності видів в екології використовується ряд термінів. До екофактору, відношення якого характеризує вплив на живий організм, додаються два слова: стено (гр. стенос - вузький); еври (гр. еурос - широкий) - (табл. 4).
Приклад: розвиток ікри різних риб проходить при різних температурах. Якщо ікра лосося розвивається при температурі від 0 до 14 °С (при оптимумі 4 °С), то по відношенню до ікри жаби звичайної вона буде стенотермна, оскільки температурні межі розвитку ікри жаби - від 0 до 30 °С з оптимумом 22 °С.
Таблиця 4
Характеристика амплітуди толерантності видів
| Екологічний фактор | Стенобіонтні | Еврибіонтні |
| Температура | стенотермні | евритермні |
| Тиск | стенобарні | еврибарні |
| Вода | стеногідричні | евригідричні |
| Глибина | стенобатні | еврибатні |
| Їжа | стенофагні | еврифагні |
| Солоність | стеногалінні | евригалінні |
| Місце проживання | стеноойкні | евриойкні |
Температура на земній поверхні залежить від географічної широти й висоти над рівнем моря. Крім того, вона змінюється з порами року.
Живі організми можуть існувати тільки в певних темпера-турних умовах. При високій температурі близько 100°С руйну-ються білки організму, а при низькій уповільнюється, а потім і припиняється обмін речовин. В залежності від температурного режиму виділяють чотири основні кліматичні зони: тропічний пояс (температура не нижче 15 - 20°С), субтропічний пояс (най-нижча температура +4 °С), помірний пояс (коливання темпера-тури від - 20 до +30 °С) та холодний пояс.
У зв'язку з цим у тварин і рослин є різні пристосування до температурних умов. У більшості організмів процеси життє-діяльності відбуваються в межах від мінус 4 до плюс 40 - 45°С. Цим пояснюється бідність форм життя в арктичних районах і в умовах тундри. Важливим для організмів є сезонний розподіл температур. Тут виникає ціла низка пристосувань організмів - зимова сплячка, сезонні міграції тощо.
Для кожного виду характерна своя оптимальна температура та крайні межі виживання, при яких відбуваються процеси життєдіяльності. Більшість морських безхребетних дуже чут-ливі до змін температури й витримують її підвищення лише до плюс 30°С і рідко які з них - до плюс 38 °С. Це мешканці великих водойм, які не перегріваються, тому в них не виникло пристосу-вань до виживання при високій температурі. Значно ширший діапазон витривалості до змін температури у мешканців малих прісних водойм. Вони можуть витримувати як промерзання, так
і нагрівання до плюс 41 - 44°С.
У багатьох організмів (рослин і всіх тварин, окрім птахів і ссав-ців) температура тіла залежить від навколишнього середовища, їх називають пойкілотермними. Інтенсивність життєдіяльності та темпи розвитку у них залежать від зовнішньої температури. Знання інтенсивності розвитку тих чи інших організмів при різних температурах важливе для проведення заходів по боротьбі з комахами - шкідниками сільського господарства або переносни-ками збудників хвороб.
Квітки багатьох рослин на ніч і в негоду закриваються, що захищає їх від переохолодження.
Під час інтенсивного руху (наприклад, при літанні) у комах тимчасово підвищується температура тіла на декілька градусів. Однак у стані спокою вона зрівнюється з температурою навко-лишнього середовища. У деяких гуртових комах (наприклад, бджіл) існує спосіб підтримання температури шляхом колектив-ної терморегуляції. Бджолина сім'я, в якій налічується кілька тисяч особин, виділяє стільки тепла, що у вулику встановлюється стала температура 34 - 35°С, що необхідно для розвитку личинок.
Найдосконаліша терморегуляція з'явилась лише у вищих хребетних - птахів і ссавців, забезпечивши їм широке розселення у всіх кліматичних поясах. Вони називаються гомойотермними організмами.
У гомойотермних тварин терморегуляція здійснюється змі-ною окисно-відновних процесів, які продукують тепло, також пристосуваннями для охолодження. У більшості ссавців охоло-дження досягається в результаті випаровування поту з поверхні шкіри та вологи із слизових оболонок. Волосяний покрив у ссав-ців і пір'я у птахів, підшкірні запаси жиру також забезпечують терморегуляцію. У гніздах тварин (норах, лігві) створюється сво-єрідний, найсприятливіший для них мікроклімат.
ЦІКАВО
Риби обживають водойми з різним температурним режимом: у гарячих джерелах Каліфорнії живе рибка луканія; риба далія мешкає в промерзлих водоймах Чукотки і Аляски; карась, вмерзаючи в лід, залишається живим. За здатністю витримувати коливання температур риб поділяють на евритерм-них, що можуть жити в широкому інтервалі температур (щука, карась, короп) і стенотермних, пристосованих до життя у вузькому температурному інтервалі - риби тропічних і полярних зон та риби значних глибин, де температура мало змінюється.
Вологість. Без води життя неможливе. Вміст води у живих клітинах в середньому становить 80-92 %. Джерелом води є опади та ґрунтові води. Тут лімітуючим фактором є кількість опадів. Вона визначає навіть тип екосистем. При опадах менш ніж 250 мм на рік формуються пустельні екосистеми; 250-750 мм - степові, лісостепові та савани; 750-1250 мм - субтропічні ліси, а більш як 1250 мм - вологі тропічні ліси.
Особливо чутливими до зміни вологості і кількості води є рос-лини. За потребою у воді виділяють такі екологічні групи рослин:
- гідатофіти, які живуть тільки у воді, вони цілком або майже повністю занурені у воду та гідрофіти, які частково зану-рені у воду. Без води вони швидко гинуть;
- гігрофіти - це наземні рослини, які можуть існувати в умо-вах підвищеної вологості повітря і на вологих ґрунтах;
- мезофіти, які ростуть в умовах помірної вологи, витри-мують нетривалу і не дуже сильну посуху. Ці рослини є досить чисельні і поширені;
- ксерофіти, які можуть переносити тривалу посуху, перебу-ваючи в активному стані завдяки здатності регулювати водний обмін, наприклад, рослини пустель, тощо.
У мешканців посушливих місць виробився ряд пристосу-вань для існування в умовах водного дефіциту. Рослини степів і пустель (ксерофіти) можуть мати видозмінені листки (колючки у кактуса) або бути безлистими (саксаул). Деякі мають дуже гли-бокі корені (наприклад, верблюжа (колючка - до 16 м). У ковили листки складаються в трубочки, продихами всередину, чим зменшується випаровування. Випаровуванню перешкоджають і такі пристосування, як щільна кутикула, восковий наліт, виро-сти шкірки волоски на поверхні листків.
Рослини-ефемери мають короткий вегетаційний період (зірочки маленькі, тюльпан, мак, шафран), весною до настання посушливого періоду встигають відквітувати, утворити насіння, запасти поживні речовини в цибулинах, кореневищах, бульбах.
Тварини також мають певні пристосування для добування та утримування води. У водних тварин і ссавців відсутній дефіцит води і тому основний продукт азотистого обміну виводиться з організму у вигляді водного розчину сечовини. Навпаки, біль-шість наземних тварин економлять воду, виводячи азот у вигляді нерозчинної у воді сечової кислоти.
Більшість тварин, мешканців пустель, може жити без води, отримуючи її з їжею або на безводний період відкладаючи багато жиру, при окисненні якого в організмі утворюються молекули води; деякі впадають у літню сплячку (гризуни, черепахи). Багато мешканців пустель рятуються і від спеки та втрати вологи, хова-ючись на день у норах. Великі ссавці пустель (сайгак, кулан) можуть здійснювати міграції на далекі відстані в пошуках води.
Спостерігаючи за птахами, польський зоолог К.А. Глогер (1838) написав книгу „Зміни у птахів під впливом клімату”, у якій він говорить про те що види тварин, які мешкають у теплих та воло-гих зонах, мають інтенсивнішу пігментацію тіла (найчастіше чорну або темно- коричневу), ніж мешканці холодних та сухих областей. Ця закономірність отримала назву „Правило Глогера”, яке поширюється на 80 % досліджених птахів та ссавців, а серед комах - на багатьох метеликів.
ЦІКАВО
Відкладання яєць у комах відбувається лише при певній вологості повітря. Комарі не кусаються, якщо відносна вологість повітря нижча за 40 %. Верблюд і одежна міль отриму-ють воду метаболічним шляхом, окислюючи жири свого тіла.
Світло - один з найважливіших факторів, з яким пов'язане все життя на Землі. У спектрі сонячного світла виділяють три біологічно нерівнозначні зони: ультрафіолетову, видиму та інфрачервону. Ультрафіолетові промені згубні для всього живого. Життя на поверхні Землі можливе завдяки озоно-вому екрану, який не пропускає основну масу цих променів. Невеликі їх кількості, що досягають поверхні землі, необхідні для життя, з ними, зокрема, пов'язаний синтез кальциферолів в організмі людини і тварин.
Видимі промені особливо необхідні для життя. Вони використовуються зеленими рослинами для фотосинтезу. Більшість тварин добре розрізняють ці промені, без них неможливе орієнтування в просторі за допомогою зору. За відношенням до світла тварин поділяють на „нічні” (активні вночі) та „денні” (активні у світлу частину доби). Розвиток кольорового зору спричинив у процесі природного добору формування різного забарвлення у тварин, що часто має захисне значення квіток, що приваблю-ють до себе комах-запилювачів.
Інфрачервоні промені найбагатші тепловою енергією. Поглинаючись тканинами тварин і рослин, вони викликають їх нагрівання. Із ними пов'язана інтенсивність фізіологічних про-цесів у рослин і пойкілотермних тварин.
Характер освітлення має добову й сезонну періодичність. У зв'язку з цим у різних видів тварин виникла пристосова-ність до активного життя в різний час доби. Майже всі фізі-ологічні процеси у рослин і тварин мають добовий ритм. Люди це відчувають при швидкому переміщенні (напри-клад, на літаку) із одного часового поясу в інші. Реакція тва-рин і рослин на тривалість світлового дня та ночі відома як фотоперіодизм.
Тривалість світлового дня змінюється з порами року, тому багато видів тварин і рослин мають річні цикли розвитку. Тривалість світлового дня впливає на статеву функцію багатьох тварин. Збільшення світлового дня навесні стимулює діяль-ність статевих залоз, а зменшення його восени веде до зату-хання їхньої функції. Скорочення дня передує похолоданню, тому в процесі еволюції органічного світу для багатьох видів зменшення світлового дня стало біологічним сигналом під-готовки до зими (підготовка до перельоту у птахів, линяння, нагромадження жиру, формування стадій спокою).
Різна тривалість дня на різних широтах обумовила появу рос-лин, що розквітають лише при короткому дні (короткоденні) на півночі й лише при довгому (довгоденні) на півдні. Багато рос-лин короткого дня, перенесені в умови довгого дня, утворюють велику вегетативну масу, але не завершують розвитку і не пере-ходять до плодоношення.
Знання закономірностей фотоперіодизму використову-ються в сільськогосподарській практиці, наприклад, при цілорічному вирощуванні овочів і декоративних рослин в умовах штучного освітлення. На птахофермах штучним освіт-ленням збільшують „світловий день” і досягають збільшення яйценосності.
Світло відіграє вирішальну роль у життєдіяльності рослин, оскільки рослини синтезують органічні речовини з неорганіч-них, використовуючи світлову енергію сонця. Одним з видів кон-куренції у рослин є конкуренція за світло. У тварин зміна трива-лості дня викликає зміну поведінки. Так, деякі птахи готуються до перельотів, інші приступають до линьки, розмноження тощо. Середня ефективність використання сонячної енергії у рослин - 1 %, максимальна продуктивність фотосинтезу за сприятливих умов - 310%.
Територія є важливим фактором у житті кожного живого орга-нізму. Всім відомо як ретельно оберігають свою територію тва-рини, ставлячи відповідні мітки. Це і зрозуміло, бо всяке життя починається з відмірювання життєвого простору, на якому меш-кають організми та виводять потомство.
5.3.
Еще по теме Принцип лімітуючих факторів:
- Класифікація екологічних факторів
- Система факторів забезпечення безпеки
- Лекція № 2 Тема: Патогенна дія факторів зовнішнього середовища.
- 5.5. Адаптація організму до змін факторів зовнішнього середовища
- ТЕМА 2.4. ТЕОРІЯ РИНКОВИХ СТРУКТУР ТА РИНОК ФАКТОРІВ ВИРОБНИЦТВА
- § 2. Соотношение принципов уголовного законодательства с принципами уголовной ответственности, принципами уголовного права, принципами уголовноправовой политики и принципами кодификации уголовноправовых норм
- Серед важливих факторів умов проживання людини є забруднення природного середовища.
- Ринок факторів виробництва ірозподіл доходів. Домогосподарства як постачальники ресурсів виробництва
- 4.Принципы успешной коммуникационной деятельности: принцип самоутверждения адресанта в глазах адресата.
- Место принципа неотвратимости юридической ответе і венпости в системе принципов законности
- Место принципа неотвратимости юридической ответственности в системе принципов законности
- 3.Принципы успешной коммуникационной деятельности: принцип вежливости Р.Лакоффа и Дж.Лича.
- Вариационный принцип Гамильтона (Принцип наименьшего действия). Уравнение движения Лагранжа 2 рода как следствие ПНД
- Вариационный принцип Гамильтона (Принцип наименьшего действия). Уравнение движения Лагранжа 2 рода как следствие ПНД
- 2. Принципы успешной коммуникационной деятельности: принцип кооперативного сотрудничества П.Грайса.
- 22. Принципы соотношения звука и буквы. Слоговой принцип русской графики
- 41 Определение принципа права. Классификация принципов права: критерии и виды.
- 1.4. Головні екологічні закони, правила і принципи
- Принципы гражданского права